39.2 RESPIRATORY GAS EXCHANGE
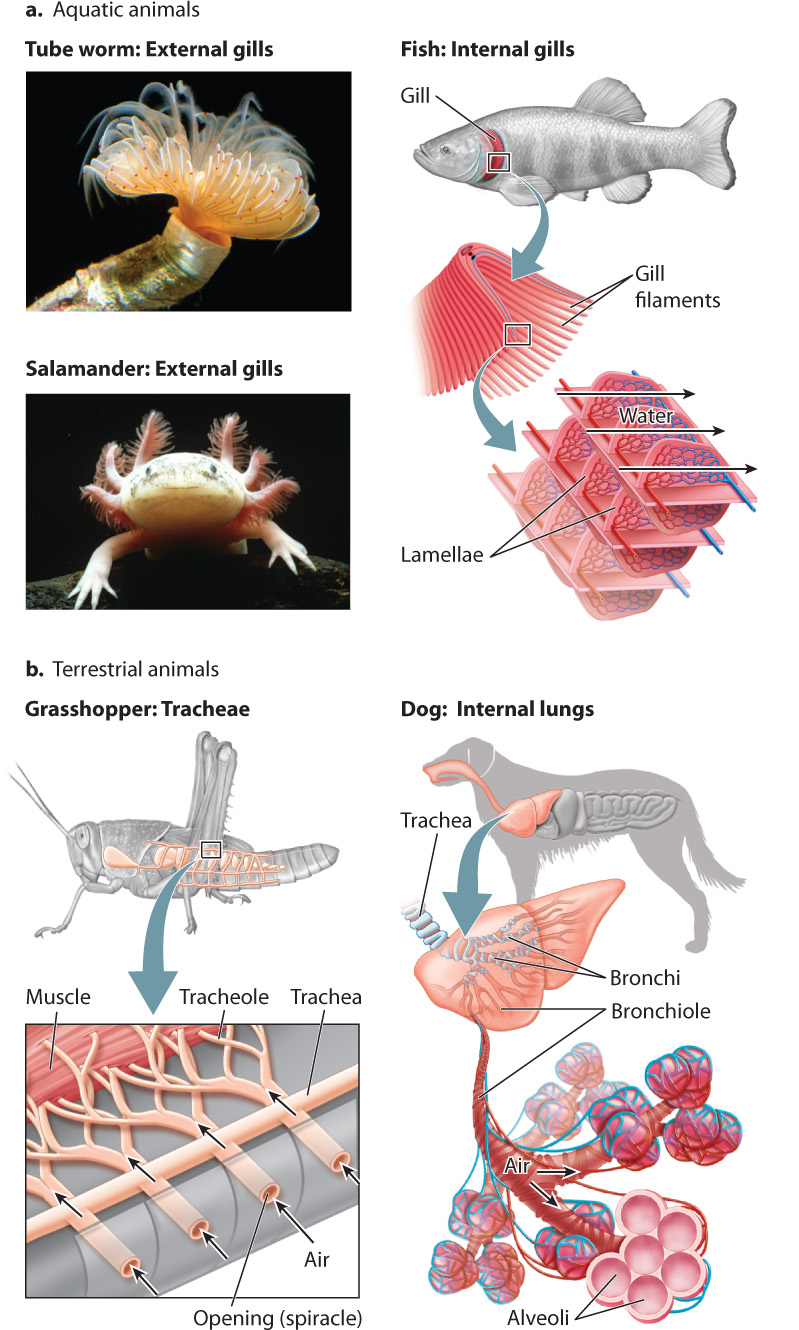
All animals obtain O2 from the surrounding air or water. Aquatic animals like crabs, aquatic salamanders, and fish that take in O2 from water breathe through gills, highly folded delicate structures that facilitate gas exchange with the surrounding water (Fig. 39.4a). Some animals, such as frogs and salamanders, also breathe through their skin. In contrast, many terrestrial animals, such as reptiles, birds, and mammals, have internal lungs for gas exchange (Fig. 39.4b). Instead of lungs, terrestrial insects evolved a system of air tubes called tracheae that branch from openings along their abdominal surface into smaller airways. These smaller airways, termed tracheoles, supply air directly to the cells within their body.
39-4
For both aquatic and terrestrial animals, ventilation supplies O2 to gas exchange surfaces, and then O2 crosses the surface through diffusion. Recall that diffusion is much slower than the bulk flow of ventilation. How can the rate of diffusion through the gas exchange surface in lungs or gills keep up with the rate of O2 supplied by ventilation? The gas exchange surface must have a large surface area and be extremely thin. The respiratory surfaces of more complex and active animals are highly folded, creating a large surface area within a small space. These surfaces are also only one or two cell layers thick, providing a diffusion distance of as little as 1 to 2 μm. In fact, this short distance is all that separates the air you breathe into your lungs from the bloodstream into which O2 diffuses.
39.2.1 Many aquatic animals breathe through gills.
Most large aquatic animals (with the notable exception of marine mammals like dolphins and whales) use gills to breathe O2-containing water. Some sessile invertebrates, including tube worms and sea urchins, as well as nudibranch mollusks and aquatic amphibians, have external gills (Fig. 39.4a). However, because external gills are easily damaged or may be eaten by predators, most mollusks, crustaceans, and fishes have evolved internal gills within protective cavities. Whereas the external gills of invertebrates often rely on the natural motions of the water to move water past the gills, invertebrates with internal gills often have cilia that direct water over the gill’s surface.
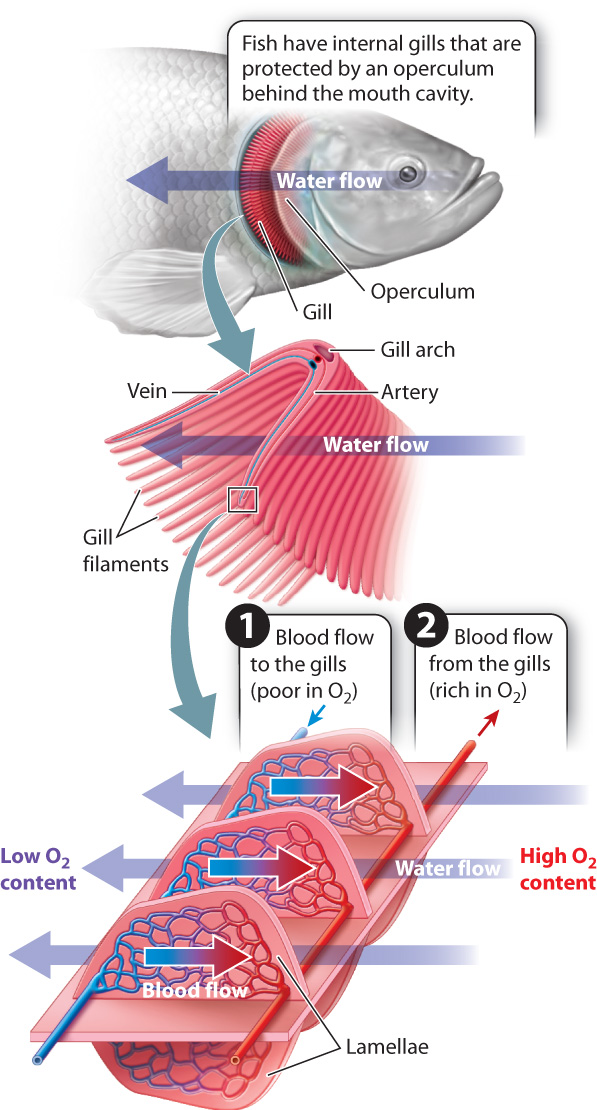
Fish need sufficient O2 to meet the energy demands of swimming. As a result, they actively pump water through their mouth and over the gills, which are located in a chamber behind the mouth cavity (Fig. 39.5). The effort required to ventilate the gills could be high because of the density and viscosity of water, but fish greatly reduce the energy cost of moving water by maintaining a continuous, unidirectional flow of water past their gills. In bony fishes, a protective flap overlying the gills, called the operculum, expands laterally to draw water over the gills while the mouth is refilling before its next pumping cycle.
The gills of fishes typically consist of a series of gill arches located on either side of the animal behind the mouth cavity and, in bony fishes, beneath the operculum. Each gill arch consists of two stacked rows of flat leaf-shaped structures called gill filaments. Numerous lamellae, thin sheetlike structures, are evenly but tightly spaced along the length of each gill filament and extend upward from the filament’s surface. A series of blood vessels brings 02-poor blood from the heart to the lamellar surfaces. The lamellae are composed of flattened epithelial cells and are extremely thin, so the water passing outside and blood passing inside the lamellae are separated by only about 1 to 2 μm. The lamellae give the gills an enormous surface area for their size.
The lamellae are oriented so that the blood flowing through them in a capillary network moves in a direction opposite to the flow of water past the gills (Fig. 39.5). This type of organization, in which fluids with different properties move in opposite directions, is an efficient way to exchange properties between the two fluids. The property can be a chemical property, such as 02 concentration, or a physical property, such as heat.
39-5
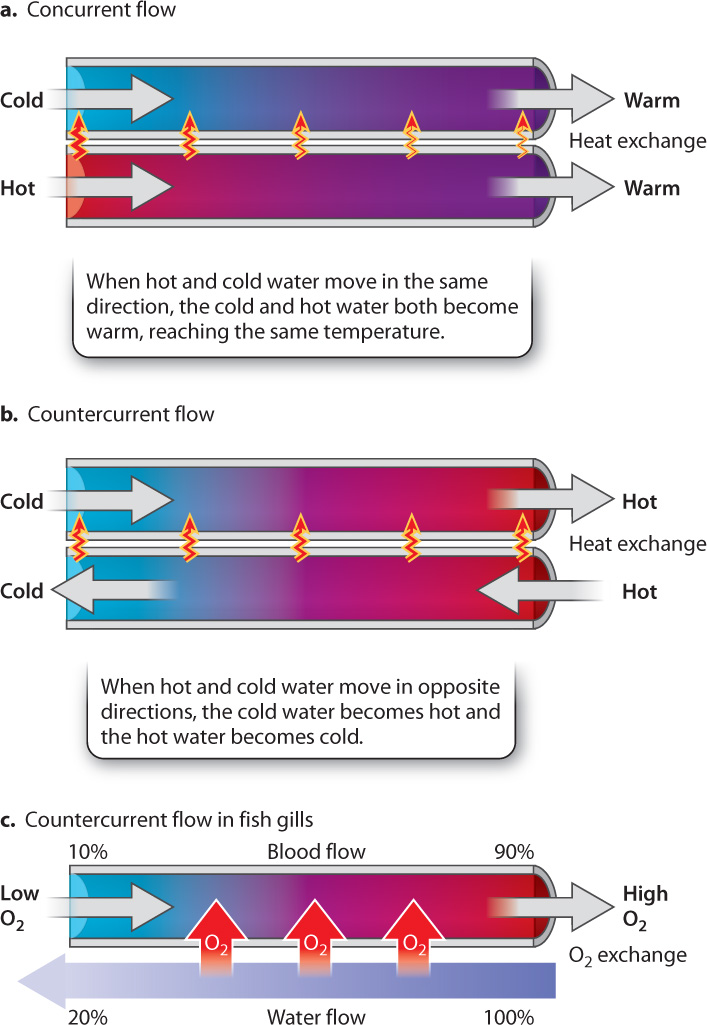
Consider for a moment two tubes right next to each other, one carrying hot water and the other carrying cold water (Fig. 39.6). If we want to make the hot water cold and the cold water hot, the efficiency of heat transfer between the two tubes depends on the direction of flow. When the water in the tubes flows in the same direction (concurrent flow), the hot water gets colder and the cold water gets hotter since heat is transferred between the two tubes (Fig. 39.6a). Given enough distance, the water will be at an intermediate (warm) temperature at the end of the tubes, reaching the same average temperature in both tubes.
Now consider what happens when the water in the two tubes flows in opposite directions (countercurrent flow). In this case, the cold water encounters increasingly hot temperatures and, given enough distance, becomes nearly as hot as the hot water entering the other tube (Fig 39.6b). Similarly, the hot water encounters increasingly cold temperatures, and becomes nearly as cold as the cold water entering the other tube. With countercurrent flow, the two essentially exchange properties (heat, in this example).
This mechanism, known as countercurrent exchange, is used widely in nature. As a result of countercurrent exchange, fish gills can extract nearly all the O2 in the water that passes over them (Fig. 39.6c). At the same time, CO2 readily diffuses out of the blood vessels and into the water that leaves the gill chamber. A set of blood vessels carries the O2-rich blood away from the gills to supply the fish’s body.
39.2.2 Insects breathe air through tracheae.
With the evolution of life on land, animals that breathe air were able to increase their uptake of O2 and achieve higher rates of metabolism. Oxygen uptake increased for the following reasons:
39-6
- The O2 content of air is typically 50 times greater than that of a similar volume of water. Fresh air contains approximately 210 ml O2 per liter of air (21%), whereas the concentration of O2 in well-mixed fresh or salt water rarely exceeds 4 ml O2 per liter of water (0.4%).
- Oxygen diffuses about 8000 times faster in air than in water. For this reason, the O2 content can be close to zero a few centimeters beneath the surface of a stagnant pond.
- Water is 800 times denser and 50 times more viscous than air. Thus, it requires more energy to pump water than to pump air past a gas exchange surface.
Because of their small size, insects can employ a direct pathway of air transport that gets air right to their tissues (see Fig. 39.4b). There is no respiratory surface. In contrast to terrestrial vertebrates, insects rely on a two-step process of ventilation and diffusion to supply their cells with O2 and eliminate CO2. First, air enters an insect through openings, called spiracles, along either side of its abdomen. The spiracles can be opened or closed to limit water loss and regulate O2 delivery, much like the leaf stomata of plants (Chapter 29). Inside the insect body, air is ventilated through a branching series of air tubes—the tracheae and tracheoles—directly to the cells. Second, diffusion occurs at the cell: O2 supplied by the fine airways diffuses into the cells, and CO2 diffuses out and is eliminated through the insect’s tracheae.
The mitochondria of the flight muscles and other metabolically active tissues of insects are located within a few micrometers of tracheole airways. Because O2 and CO2 diffuse rapidly in air, the tracheal system of insects delivers O2 to cellular mitochondria at high rates. Insects also have an air sac system connected to their tracheae. The air sacs act like bellows to pump air through the tracheae, speeding the movement of air to tissues so that gas exchange is faster.
39.2.3 Terrestrial vertebrates breathe by tidal ventilation of internal lungs.
Unlike fish gills, the lungs of most land vertebrates inflate and deflate to move fresh air with O2 into the lungs and expire stale air with CO2 out of the lungs. The low density and viscosity of air enables these animals to breathe by tidal ventilation without expending too much energy. In tidal respiration, air is drawn into the lungs during inhalation and then moved out during exhalation (Fig. 39.7a).
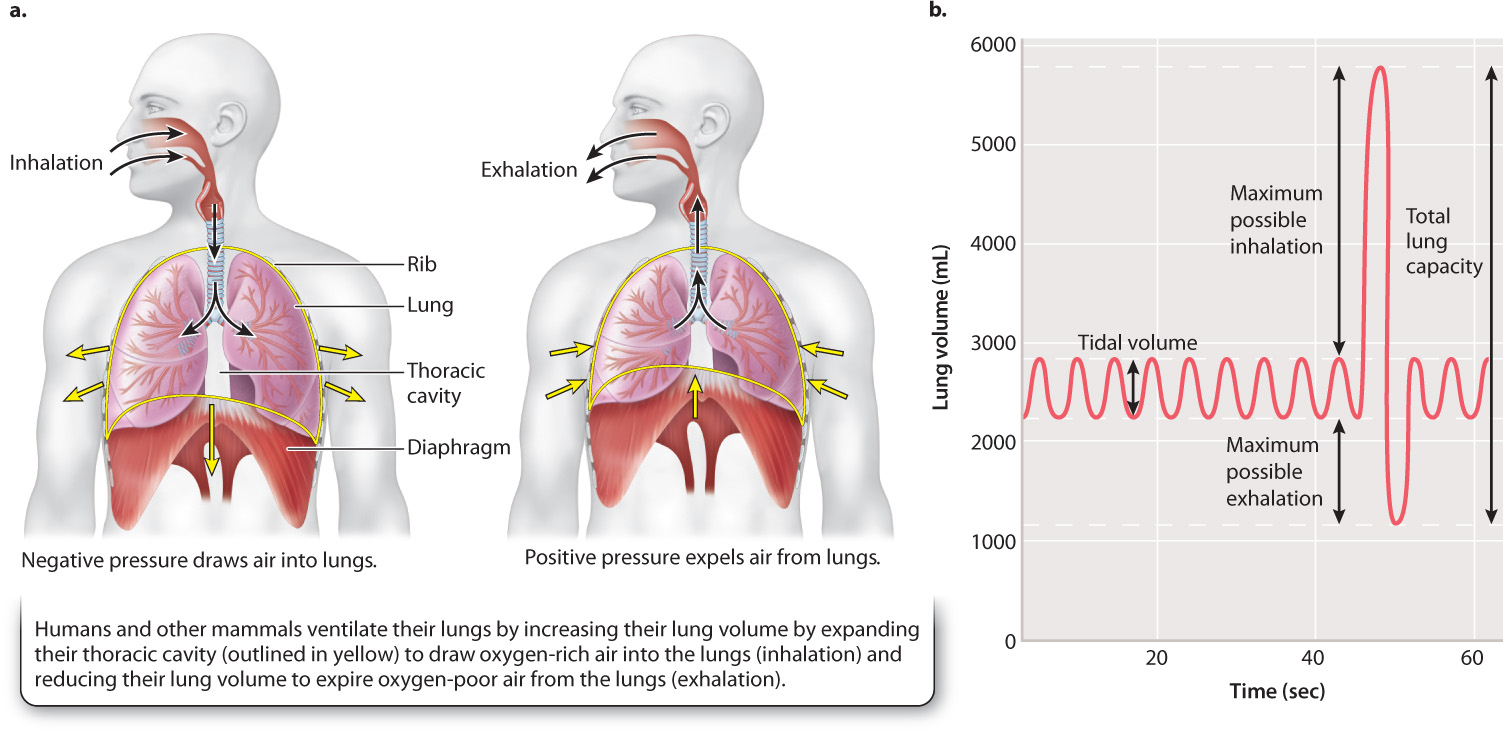
Mammals and reptiles expand their thoracic cavity to draw air inside their lungs on inhalation. The expansion of the lungs causes the air pressure inside lungs to become lower than the air pressure outside the lungs. The resulting negative pressure draws air into the lungs. By contrast, amphibians inflate their lungs by pressure produced by their mouth cavity. In most animals, exhalation is passively driven by elastic recoil of tissues that were previously stretched during inhalation. The contraction of the lungs causes the air pressure inside the lungs to become higher than the air pressure outside the lungs. The resulting positive pressure forces air out of the lungs.
In mammals, inhalation during normal, relaxed breathing is driven by contraction of the diaphragm, a domed sheet of muscle located at the base of the lungs that separates the thoracic and abdominal cavities (Fig. 39.7a). Exhalation occurs passively by elastic recoil of the lungs and chest wall. During exercise, other muscles come into play to assist with inhalation and exhalation. For example, intercostal muscles, which are attached to adjacent pairs of ribs, assist the diaphragm by elevating the ribs on inhalation and depressing them during exhalation. The action of the intercostal muscles helps to produce larger changes in the volume of the thoracic cavity, increasing the negative pressure that draws air into the lungs during inhalation and assisting elastic recoil of the lungs and chest wall to pump air out of the lungs during exhalation.
39-7
At rest, we inhale and exhale about 0.5 liter of air every cycle (Fig. 39.7b). This amount represents the tidal volume of the lungs. With a breathing frequency of 12 breaths per minute, the ventilation rate (breathing frequency × tidal volume) is 6 liters per minute. When more O2 is needed during exercise, both breathing frequency and tidal volume increase to elevate ventilation rate.
The fresh air inhaled during tidal breathing mixes with O2-depleted stale air that remains in the airways after exhalation. As a result, the pO2 in the lung (approximately 100 mmHg in humans) is lower than the pO2 of freshly inhaled air (approximately 160 mmHg at sea level). Consequently, the fraction of O2 that can be extracted is lower than the fraction that can be extracted by the countercurrent flow of fish gills, which can reach 90% or more. Typically, mammalian lungs extract less than 25% of the O2 in the air, and reptile and amphibian lungs extract even less. Nevertheless, the disadvantages of tidal respiration are offset by the ease of ventilating the lung at high rates and the high O2 content of air.
Question Quick Check 3
pILl+v4ttdiCGG5JBYKPZz/Yhv0XDKp1WxujOY/bxLf2lGzdgb0JfqnW3c6laWuWK6BVFLe0wJX++GPPk3Vc5SDCcTrmrd5iMFAo2U+XOOJVLfmNI8z8ikD61uFJ9peM6ggNvtfz28Q2mTyx39.2.4 Mammalian lungs are well adapted for gas exchange.
Despite the limitations of tidal respiration, mammalian lungs are well adapted for breathing air. They have an enormous surface area and a short diffusion distance for gas exchange. Consequently, the lungs of mammals supply O2 quickly enough to support high metabolic rates.
Except in the case of birds (discussed below), the lungs of air-breathing vertebrates are blind-ended sacs located within the thoracic cavity. Air is taken in through the mouth and nasal passages, and then passes through the larynx, within which the vocal cords are located. (The vocal cords enable speech, song, and sound production.) Air then enters the trachea, the central airway leading to the lungs (Fig. 39.8). The trachea divides into two airways, called primary bronchi, one of which supplies each lung. The trachea and bronchi are supported by rings of cartilage that prevent them from collapsing during respiration, ensuring that airflow meets with minimal resistance. You can feel some of these cartilage rings at the front of your throat below your larynx (the part of the larynx you can feel is your Adam’s apple). The primary bronchi divide into smaller secondary bronchi and again into finer bronchioles. This branching continues until the terminal bronchioles have a diameter of less than 1 mm. The very fine bronchioles end with clusters of tiny thin-walled sacs, the alveoli, where gas exchange by diffusion takes place. As a result of this branching pattern, each lung consists of several million alveoli, providing a large surface area. Together, human lungs consist of300 to 500 million alveoli with a combined surface area as large as a tennis court—about 100 m2!
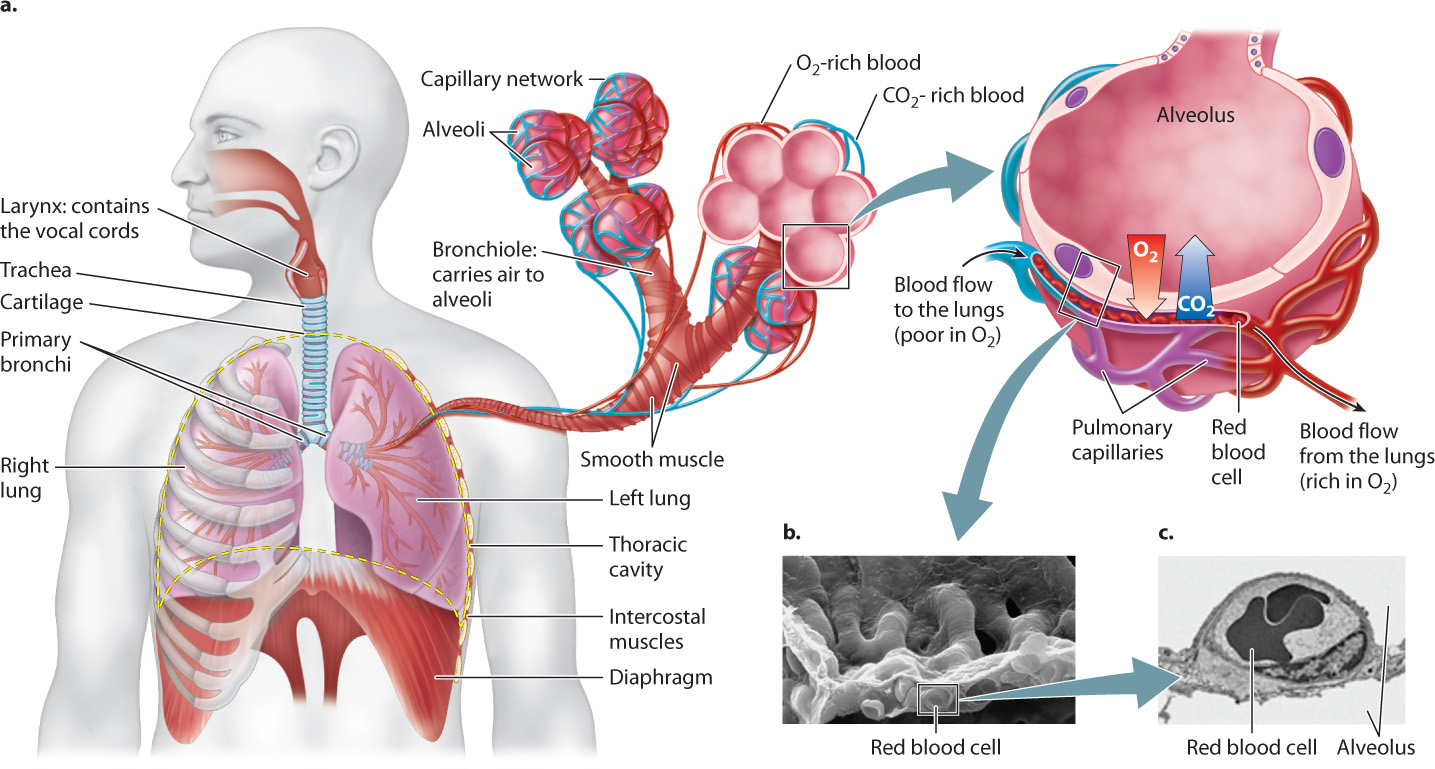
39-8
Small blood vessels, called pulmonary capillaries, supply the alveolar wall. Each alveolus is lined with thin epithelial cells in intimate contact with the endothelium of these small blood vessels (Fig. 39.8). As a result, the diffusion distance from the alveolus into the capillary is extremely short (about 2 μm), similar to that of gill lamellae and the terminal airways of bird lungs.
Keeping the surfaces of the alveoli moist is critical because moisture helps move O2 molecules from the air into solution and thus diffuse across the alveolar wall. Mucus-secreting cells lining the alveoli keep the inside surface of the alveoli coated with a fluid film. Other alveolar epithelial cells produce a compound called surfactant, which acts like soap to reduce the surface tension of the fluid film. Surface tension is a cohesive force that holds the molecules of a liquid together, causing the surface to act like an elastic membrane. Because of surface tension, it requires greater pressure to inflate a small balloon than a large one. The surfactant allows the lungs to be inflated easily at low volumes when the alveoli are partially collapsed and small. It also ensures that alveoli of different sizes inflate with similar ease, enabling more nearly uniform ventilation of the lung.
Mucus-secreting cells line all the airways of the lung not only to keep them moist but also to trap and remove foreign particles and microorganisms that an animal may breathe in with air. Beating cilia on the surface of these cells move the mucus and foreign debris out of the lungs and into the throat, where it is swallowed and digested. Smoking even one cigarette can stop the cilia’s beating for several hours or even kill them. This is one reason why smokers cough—they must clear the mucus that builds up.
39.2.5 The structure of bird lungs allow unidirectional airflow for increased oxygen uptake.
Birds extract O2 efficiently to supply the considerable energy needed for flight. An efficient O2 supply is particularly important for migratory birds that fly at high altitude where the air’s partial pressure of O2 is relatively low. The lungs of birds are unique in that they are rigid and do not inflate and deflate. As a consequence, birds do not use tidal breathing. Instead, they benefit from unidirectional airflow that enhances gas exchange by maintaining larger concentration gradients for diffusion.
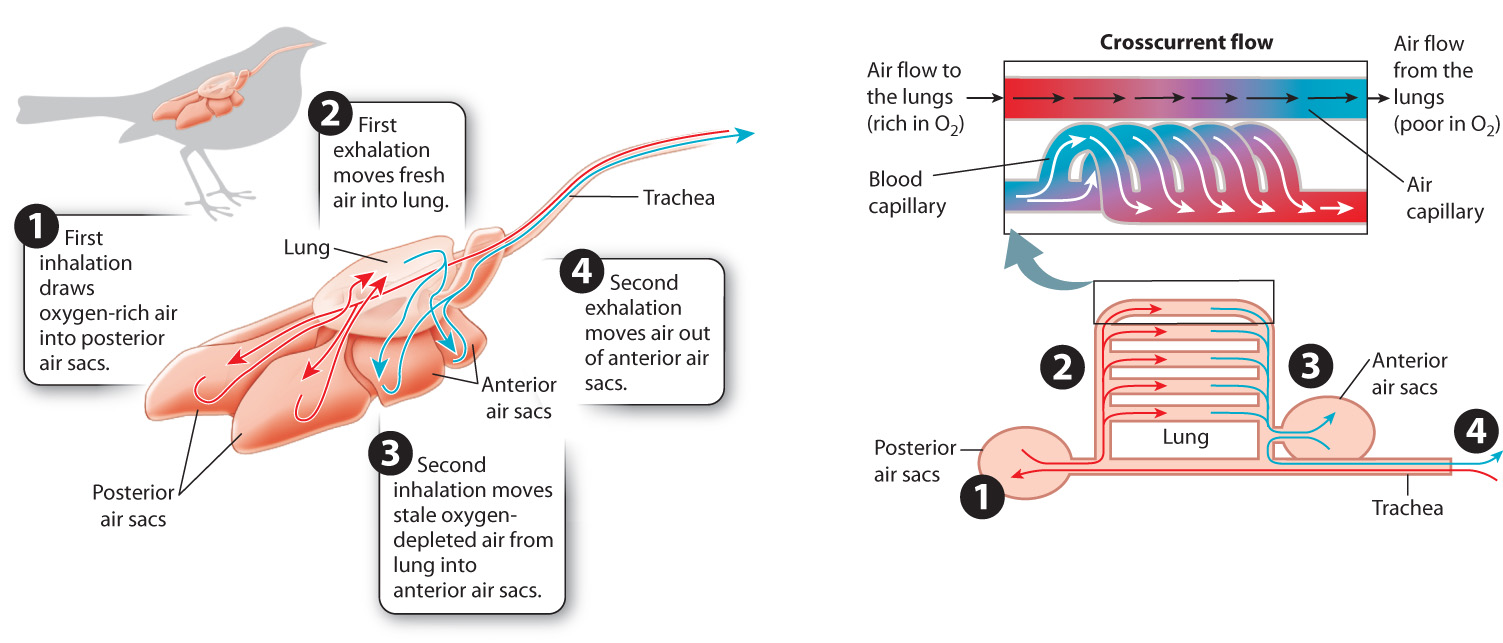
A bird’s respiratory system consists of two sets of air sacs: one located posterior to the lungs (behind them) and one located anterior to the lungs (in front of them). Air is pumped through these air sacs in a bellows-like action (Fig. 39.9). As the set of posterior air sacs expand, air is drawn from the bird’s mouth into those sacs. As the sacs are then compressed by surrounding muscles acting on the skeleton, air is pumped through the bird’s rigid lungs.
39-9
The lungs consist of a series of larger air channels that branch into fine air capillaries having a large surface area. The air capillaries direct the air so that it moves crosscurrent (at 90°) to the path of blood flow through the walls of the air capillaries. This crosscurrent flow is not as efficient as the countercurrent flow of fish gills, but it enables birds to extract more O2 than mammals can.
After losing O2 and gaining CO2, the air leaves the lungs and enters the set of anterior air sacs. After fresh air is inhaled into the posterior air sacs, the stale air in the anterior air sacs is exhaled out of the bird’s trachea and mouth. As a result, birds achieve a continuous supply of fresh air into the lung during both inhalation and exhalation. In contrast to other air-breathing vertebrates, therefore, birds move air through their respiratory system in two cycles of ventilation.
39.2.6 Voluntary and involuntary mechanisms control breathing.
Because an animal’s need for O2 varies with activity level, animals adjust their respiratory rate to meet their cells’ changing demand for O2. Respiration is a unique physiological process in that it is controlled by both the voluntary and involuntary components of the nervous system (Chapter 35). In sleep, breathing is maintained at a resting rate by the involuntary part of the nervous system. Indeed, in most circumstances, breathing is controlled unconsciously.
The regulation of blood O2 levels is a key example of homeostasis, as we saw in the case of core body temperature (Chapter 35) and blood glucose levels (Chapter 38). Recall that homeostasis often depends on sensors that monitor the levels of the chemical being regulated. In the case of breathing, the sensors are chemoreceptors located within the brainstem and in sensory structures called the carotid and aortic bodies (Fig. 39.10). The carotid bodies sense O2 and proton (H+) concentrations of the blood going to the brain, and the aortic bodies monitor their levels in blood moving to the body. In contrast, chemoreceptors in the brainstem sense CO2 and H+ concentrations. The most important factor in the control of breathing is the amount of CO2 in the blood. If the concentration of CO2 in the blood is too high, chemoreceptors in the brainstem stimulate motor neurons that activate the respiratory muscles to contract more strongly or more frequently. Stronger or faster breathing rids the blood of excess CO2 and increases the supply of O2 to the body.
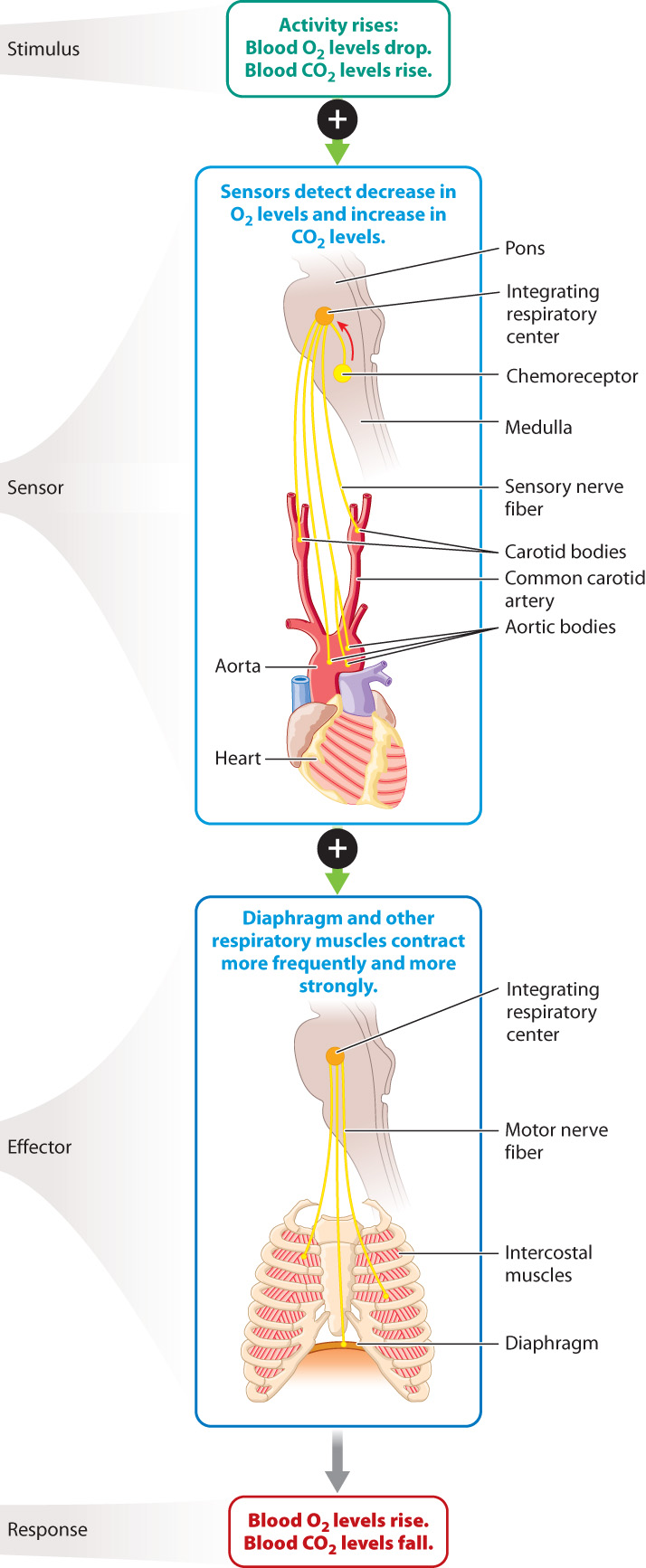
Breathing can also be controlled voluntarily. It is a simple matter to voluntarily hold your breath. Holding the breath makes it possible for humans and marine mammals to dive under water; it is also critical to the production of speech, song, and sound for communication. Sound is produced by voluntarily adjusting the magnitude and rate of airflow over the vocal cords of mammals, the syrinx of songbirds, or the glottal folds of calling amphibians, such as some toads and frogs.
39-10