40.4 DIGESTION AND ABSORPTION OF FOOD
After animals obtain food, they break it down through the process of digestion. Digestion requires that the food be isolated in a specialized compartment so that it can be broken down chemically without damage to other cellular organelles or structures of the body. Organisms on different branches of the tree of life have evolved very different means of isolating and digesting food. The process in single-celled protists, which obtain food particles by phagocytosis, is intracellular digestion, in which food is broken down within cells. Lysosomes containing hydrolytic enzymes fuse with vacuoles containing food particles, so the enzymes are mixed with the food (Chapter 5). Following their chemical breakdown, the food products of intracellular digestion are absorbed into the cell for use. In most animals, however, the process is extracellular digestion, in which food is isolated and broken down outside a cell, in a body compartment.
40.4.1 The digestive tract has regional specializations.
Extracellular digestion involves a specialized cavity or body compartment in which food is broken down. Some animals, like sea anemones, have an internal cavity in which digestion occurs. Cells lining this cavity secrete digestive enzymes that break down the food, allowing the smaller chemical compounds to be absorbed by the cells.
Other animals have more elaborate digestive systems that allow the transport of food by a digestive tube that runs from an animal’s mouth to its anus. Collectively, the passages that connect the mouth, digestive organs, and anus constitute an animal’s gut or digestive tract. Because the food is moved in a single direction through a tubelike gut, particular regions of the digestive tract can be specialized for different functions. These functions include storage, chemical breakdown of different kinds of food, absorption of released nutrients, and elimination of waste products.
The digestive tracts of animals are commonly divided into a foregut, midgut, and hindgut (Fig. 40.13). Most animals’ digestive systems have the same basic plan, though some animals have also evolved specialized structures. The foregut includes the mouth, esophagus, and stomach or crop, which serves as an initial storage and digestive chamber. Next is the midgut, which includes the small intestine, where the remainder of digestion and most nutrient absorption takes place. Here, specialized organs secrete enzymes and other chemicals that aid in the breakdown of particular macromolecules, such as fats and carbohydrates. Finally, the digested material reaches the hindgut, which includes the large intestine and rectum. Within the hindgut, water and minerals are reabsorbed, leaving the waste products, or feces, which are stored in the rectum until being eliminated from the body. Elimination of the feces, or defecation, involves relaxation of an anal sphincter combined with muscular contraction of the rectum and body wall.
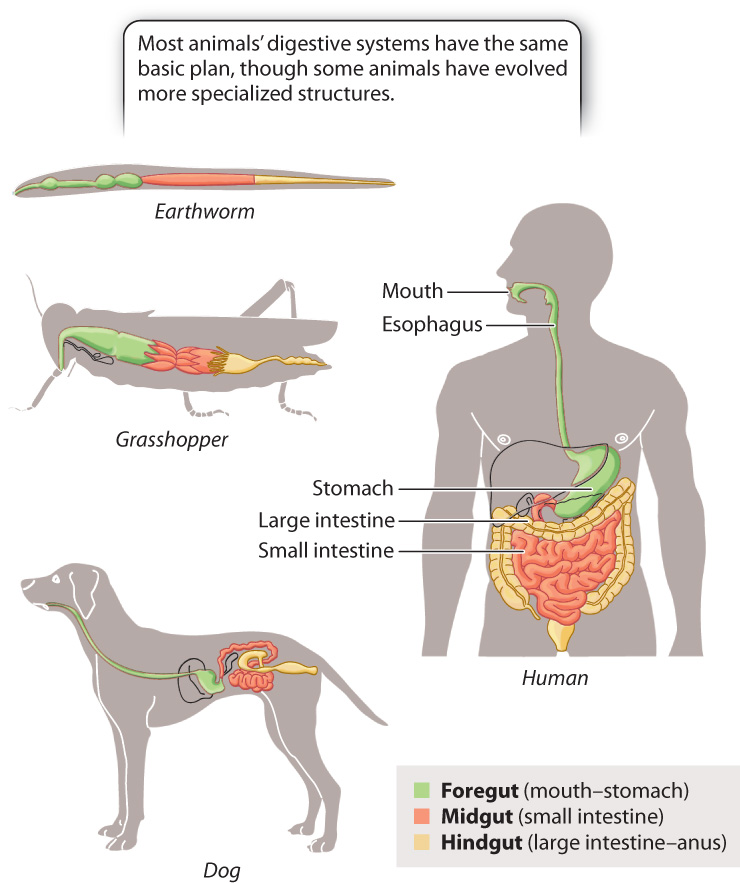
40-15
40.4.2 Digestion begins in the mouth.
Most animals jump-start digestion by breaking down food mechanically. After ingesting food, many animals manipulate the food in the mouth or buccal cavity. As we saw in the previous section, many animals break down food with the aid of jaws and teeth. Insects, which lack jaws and teeth, manipulate and break down their food with mouthparts called mandibles before it passes into their gut. In some animals, such as mammals, food entering the mouth is mixed with salivary secretions that contain amylase, an enzyme that breaks down carbohydrates, to begin the digestion of sugars and starches.
Mammals and other land vertebrates have a muscular tongue that facilitates food manipulation and transport within the mouth cavity. The tongue moves the food into position within the mouth for effective cutting and maceration into smaller pieces. When a bolus of food has been chewed and is ready to be swallowed, the tongue moves it to the rear of the mouth cavity.
Swallowing, which is controlled by the autonomic nervous system, is a complex set of motor reflexes that involves several muscles and structures in the rear of the mouth and the pharynx, the region of the throat that connects the nasal and mouth cavities (Fig. 40.14). Once initiated, swallowing reflexes occur without voluntary control. During swallowing, food is pushed through the pharynx, over the epiglottis, a flap of tissue that protects food from entering the trachea and lungs, and into the esophagus, the tube that connects the pharynx to the stomach.
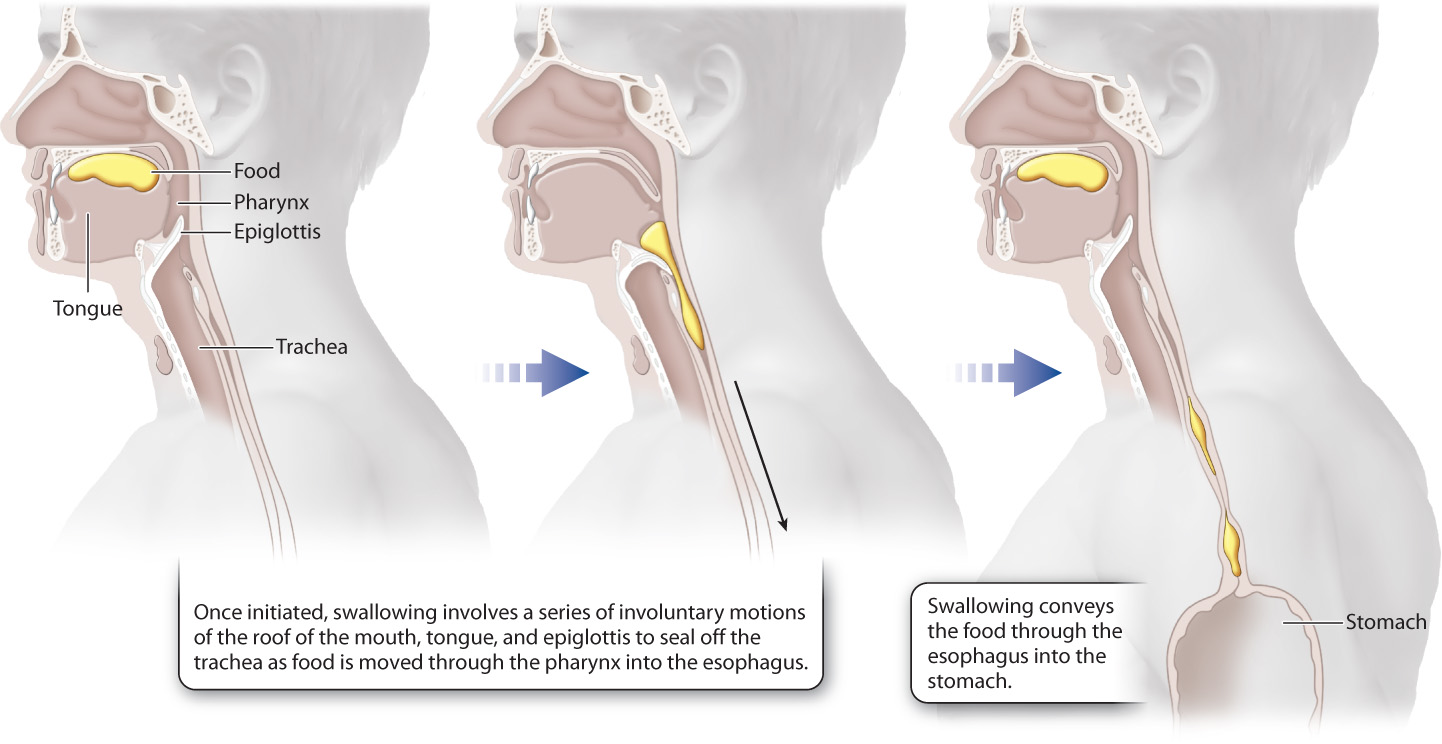
40-16
Birds, alligators, crocodiles, and earthworms break down food into smaller pieces further along their digestive tracts in the gizzard, a compartment with thick muscular walls. Birds and earthworms often ingest small rocks or sediment to help grind the food into smaller pieces within the muscular gizzard. Breaking food into smaller sizes aids the chemical digestion that follows.
Question Quick Check 4
bI0SvXlND5f6TrorpAeBBFkjYq6Ude4GIywv8DKgYwqm++cupnfMvxDwbFHgQmEOfAYAFbXUSNQWFTuXRH8lIx4iXeF9QsbRHcLQsIduY9tQZSPHNW1Pwf4Pzkuaj/ybVjiaoB9g9qhgURLxNdJp+OCQ+V4gfVJO40.4.3 The stomach is an initial storage and digestive chamber.
After being ingested and mechanically broken down, the food is transported into a stomach or crop (see Fig. 40.13). Stomachs and crops further fragment and mix the food with digestive enzymes, storing the food in a partially digested form before releasing it for further digestion and nutrient absorption, as needed by the animal.
The stomach is one of the main sites of protein breakdown. The sight, smell, and taste of food send signals to the brain that stimulate a sense of appetite, and to the stomach, stimulating the secretion of hydrochloric acid (HCl) and digestive enzymes that break down proteins into amino acids, which are then absorbed into the bloodstream. Cells secrete these digestive enzymes in an inactive form; otherwise, they would be digested themselves. After secretion, the inactive enzymes are activated by a change in their chemical structure. The primary digestive enzyme produced in the stomach is pepsin, an enzyme that breaks down proteins. Cells lining the stomach release pepsin in an inactive form called pepsinogen that is activated by the low pH of the stomach, which is in the range of pH 1–3. The low pH also aids active pepsin in cleaving bonds between amino acids of the peptides. Finally, the acidic environment and digestive enzymes of the stomach kill most pathogens that may be ingested with the food.
40-17
In addition to digestive enzymes and HCl, the cells lining the stomach secrete a peptide hormone called gastrin in response to food in the stomach (Fig. 40.15). Gastrin stimulates the cells lining the stomach to increase their production of HCl. Gastrin secreted by the stomach is absorbed within the small intestine and recirculated to the cells lining the stomach wall. There, the recirculated gastrin stimulates the stomach lining cells to increase their production of gastrin, forming a positive feedback loop (Chapter 38). The feedback loop, starting with the secretion of pepsin to the amplified secretion of gastrin, ensures that protein digestion occurs in response to the start of a meal as food enters the stomach. If the pH of the stomach becomes too low, gastrin secretion is inhibited by negative feedback control.
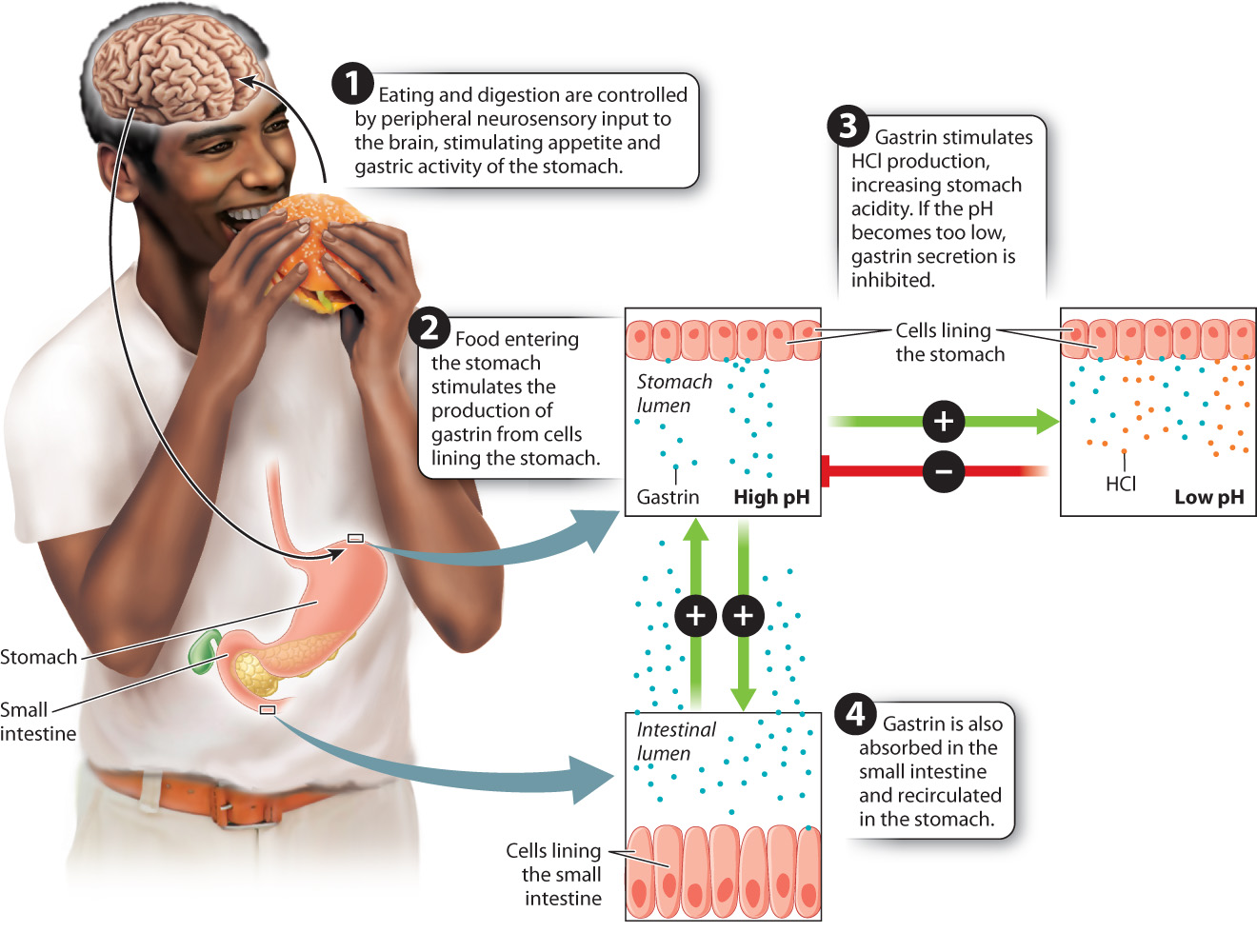
Glands in the lining of the stomach secrete mucus to protect the stomach wall from the acid its cells secrete to digest food. Sometimes, this defense can break down, resulting in an ulcer, a break or erosion in the cells lining the stomach. Although stress and oversecretion of digestive juices can cause an ulcer, Australian pathologists Barry Marshall and Robin Warren made the remarkable discovery that most ulcers result from an infection by the bacterium Heliobacter pylori, which can survive in extremely acidic environments. Ulcers produced by H. pylori are worsened by the stomach’s secretions of HCl and pepsin. The two researchers won the Nobel Prize in Physiology or Medicine in 2005 for their work.
The stomach walls contract to mix the contents of the stomach, aiding their digestion. Waves of muscular contraction, called peristalsis, move the food toward the base of the stomach. There, the pyloric sphincter, a band of muscle, opens and allows small amounts of digested food to enter the small intestine. Opening and closing of the pyloric sphincter regulates the rate at which the stomach empties, allowing time for digestion and absorption of the food products released into the small intestine.
Animals often eat large amounts of food in a short time, but it takes much longer to digest that food. The stomach serves as a storage compartment as well as a digestion compartment. In humans, it typically takes about four hours for the stomach to empty, allowing digestion and absorption of nutrients to occur between meals. In other animals, stomach emptying can take much longer. Most carnivorous animals consume large and infrequent meals, resulting in long periods of digestion and nutrient absorption compared with humans and other animals that eat more regularly. Snakes such as pythons that engulf large prey whole (Fig. 40.16) elevate their metabolic rate to high levels during digestion and spend several days digesting and absorbing the nutrients from their meal. This requires an extensive remodeling of their gut to produce new secretory and absorptive cell surfaces to digest their single large meal. These extra cells are not retained between meals because of the high energy cost of maintaining them.

40.4.4 The small intestine is specialized for nutrient absorption.
After the food enters the small intestine, protein and carbohydrate digestion continue, and fat digestion begins. The small intestine is so named because of its small diameter, but it is a long (about 20 feet or 6 m in an adult human) and therefore large organ. Because of its length, the small intestine is coiled to fit within the abdominal cavity. The small intestine is made up of three sections (Fig. 40.17). The initial section is the duodenum, into which the food enters from the stomach. Most of the remaining digestion takes place in the duodenum. The next two sections, the jejunum and ileum, carry out most nutrient absorption.
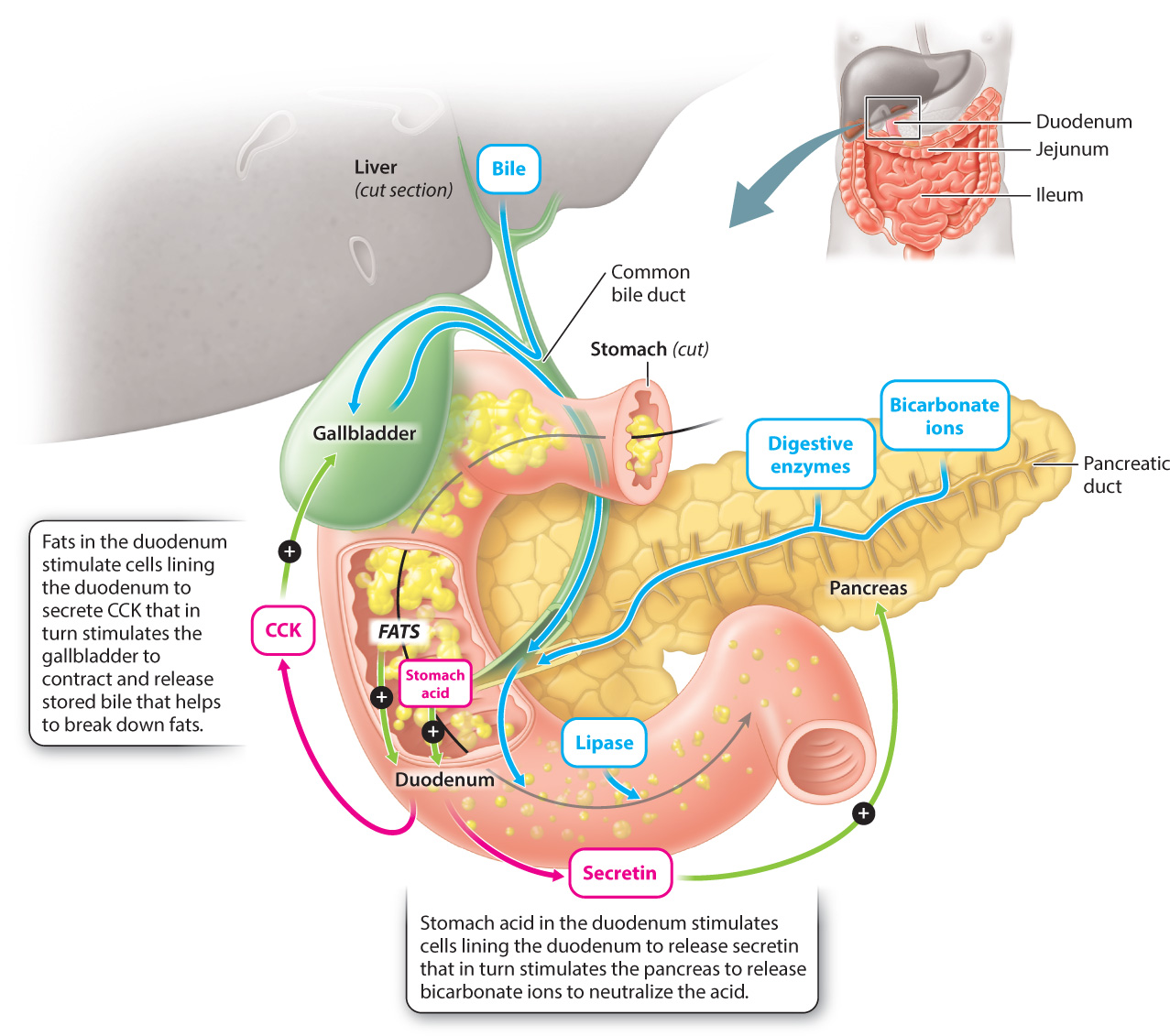
Two accessory organs, the liver and pancreas, aid in the digestion of proteins, carbohydrates, and fats in the duodenum of the small intestine. The liver produces bile, which is composed of bile salts, bile acids, and bicarbonate ions. Bile aids in fat digestion by breaking large clusters of fats into smaller lipid droplets, a process called emulsification. Bile produced by the liver is stored in the gallbladder (Fig. 40.17). When fats enter the duodenum, cells lining the duodenum release a peptide hormone called cholecystokinin (CCK), which causes the gallbladder to contract, thus releasing the bile into the duodenum. Bile in turn breaks up the fats into smaller droplets, allowing lipases, enzymes that break apart lipids such as triacylglycerol, to digest the lipids more effectively, breaking them down into free fatty acids and glycerol that can be readily absorbed across the wall of the small intestine and eventually into the bloodstream.
40-18
The pancreas is a secretory gland that lies below the stomach, posterior to the duodenum (Fig. 40.17). It functions as both an endocrine gland, secreting hormones directly into the blood as discussed in Chapter 38, and an exocrine gland, secreting substances into ducts that connect to the duodenum. The pancreas produces a variety of digestive enzymes, including lipase, which breaks down fats, and trypsin, which further breaks down proteins. Like pepsin produced by the stomach, the pancreas produces trypsin in an inactive form called trypsinogen to avoid digesting itself.
The pancreas also secretes bicarbonate ions, which neutralize the acid produced by the stomach. Bicarbonate secretion by the pancreas is stimulated by the hormone secretin (the first hormone ever to be identified), which is released by cells lining the duodenum in response to the acidic pH of the stomach contents entering the small intestine. Enzymes that break down proteins work best in acidic environments like that of the stomach, but most proteins, like those that break down carbohydrates and fats, are denatured (that is, unfolded and thus made inactive) in acidic environments. In order to digest carbohydrates and fats, the stomach acid is neutralized by the secretion of bicarbonate ions by the pancreas, which then allows the enzymes that digest carbohydrates and fat to operate.
40-19
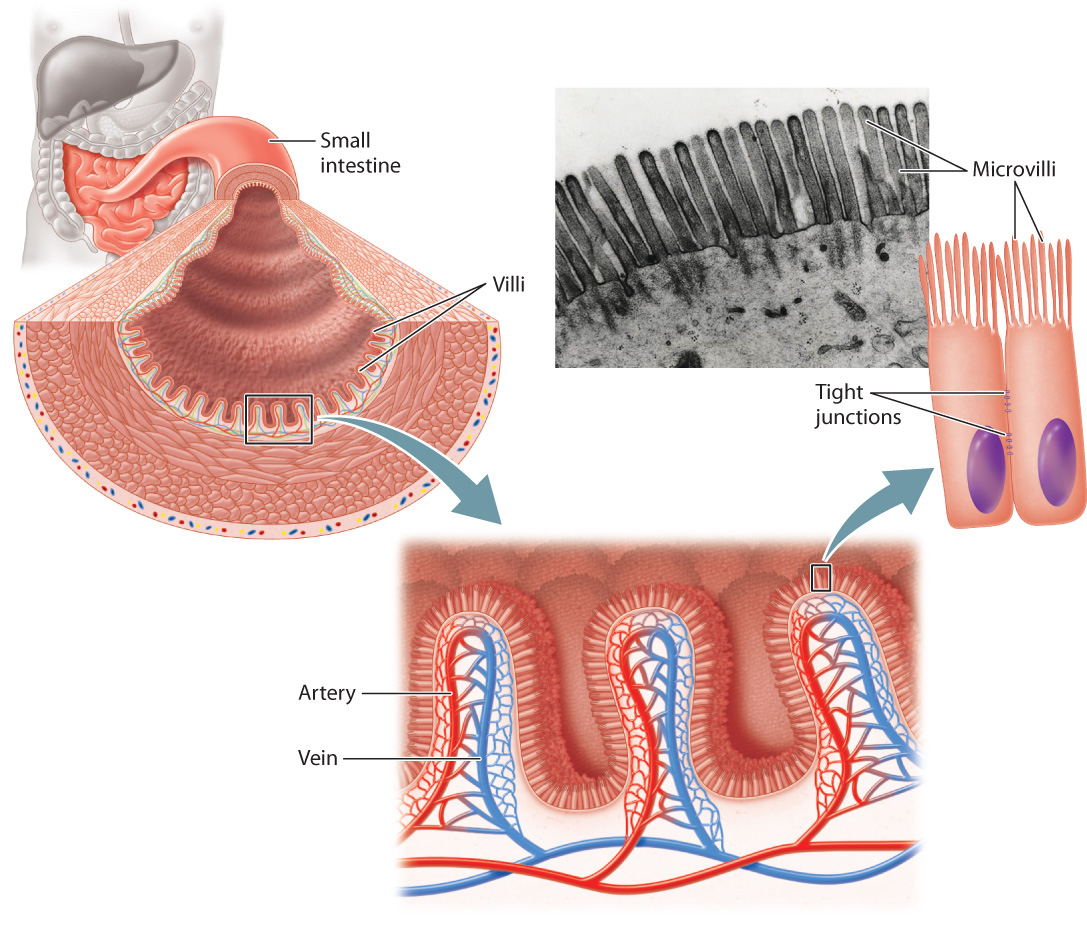
Final digestion of proteins and carbohydrates occurs in the jejunum and ileum. These two sections of the small intestine have highly folded inner surfaces, called villi, and the cells constituting the villi themselves have highly folded surfaces, called microvilli (Fig. 40.18). Together, villi and microvilli greatly increase the surface area for the absorption of nutrients. Maximizing surface area by folding into villi and microvilli is much like the way the lung maximizes surface area for gas exchange with alveoli (Chapter 39). Cells that line the small intestine are connected by tight junctions (Fig. 40.17 and Chapter 10). These junctions force the products of digestion to be absorbed across the microvilli surfaces, controlling their movement through the cell and into the bloodstream, rather than leaking between the cells.
The microvilli of the cells lining the small intestine secrete enzymes that cleave peptides into amino acids, which can then be absorbed across the plasma membrane. They also secrete enzymes that break sugars into their subunits, which can also be readily absorbed. Lactose intolerance can arise in humans if they stop producing lactase, an enzyme that normally breaks lactose into its component parts. This can be a critical problem for infants who fail to produce lactase, as milk is their principal source of nutrition. Lactose intolerance is readily treated by lactase supplements to the diet.
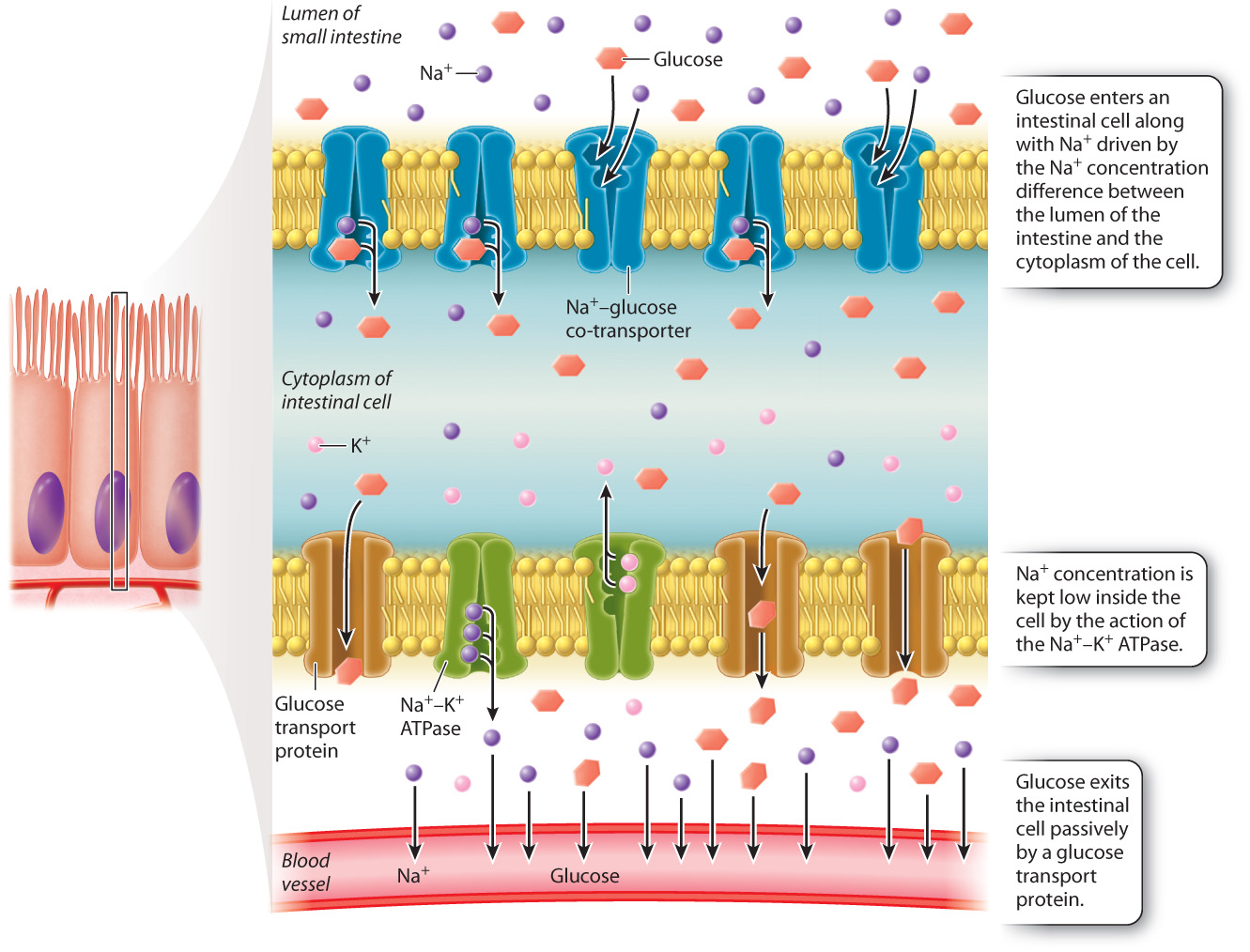
Nutrient molecules, such as glucose and amino acids, are often co-transported into cells lining the intestine with sodium ions from gut contents. The sodium ion binds to a transmembrane protein on the surface of cells lining the intestine that also binds the nutrient molecule. Absorption of the nutrient molecule into the cell is driven by the movement of the sodium ion down its concentration gradient (Fig. 40.19). The concentration of sodium ions inside the cell is kept low by the action of the sodium-potassium pump (Chapter 5). Molecules such as glucose are then moved from the intestinal cell into the blood by a pump on the basal side of the cell. Because the products of fat digestion—fatty acids and glycerol—are lipid soluble, they do not require a carrier protein for transport. Instead, they readily diffuse across the lipid plasma membrane, entering the mucosal cell, where they are further broken down and eventually transported to the bloodstream.
40-20
40.4.5 The large intestine reabsorbs water and stores waste.
After the absorption of nutrients in the small intestine, water and mineral ions are reabsorbed in the large intestine, or colon, of the hindgut (see Fig. 40.13). Peristalsis moves the contents of the small intestine into the colon at a pace that allows time for nutrient absorption in the small intestine. By the time the gut contents reach the large intestine, the nutrients have been absorbed into the body, but water and inorganic ions remain. These are absorbed in the large intestine until the contents form semisolid feces. Excess water absorption can cause constipation, whereas too little water reabsorption results in diarrhea. Diarrhea and constipation also can be caused by toxins released by microorganisms, such as the bacterium Vibrio cholera, the cause of cholera, in the food an animal eats. Feces are stored in the final segment of the colon, the rectum, until they are periodically eliminated through the anus.
As we saw in Case 5: The Human Microbiome, digestion is not a solo endeavor. Large populations of bacteria reside in the small and large intestine and help extract nutrients that the animal’s body cannot extract itself. The principle gut resident is Escherichia coli, the bacterium commonly used for research in molecular biology laboratories. In the process of nourishing themselves by aiding in the digestion of the host’s gut contents, the bacteria provide nutrients and certain vitamins, such as biotin and vitamin K, that the animal cannot produce itself. Vitamin deficiency therefore can sometimes result from prolonged antibiotic medication that kills large numbers of gut bacteria. Another by-product of bacterial action in the gut is gases such as methane and hydrogen sulfide. These are expelled when foods contain compounds that the bacteria can digest but which cannot be digested and absorbed by the host animal, as when humans eat beans. The mutual benefits to the host animal and the bacteria ensure the success of their symbiotic relationship.
40-21
40.4.6 The lining of the digestive tract is composed of distinct layers.
As we have seen, the digestive tract is not a passive tube. It secretes enzymes and other chemical compounds, absorbs nutrients, and actively moves food through the body. How does a series of tubes accomplish all of this? As shown in Fig. 40.20, the digestive tract is made up of several layers of tissue, each with a specialized function. The central space, or lumen, through which the gut contents travel is surrounded by an inner tissue layer, the mucosa, which has secretory and absorptive functions. The cells of the mucosa secrete mucus to protect the gut wall from digestive enzymes, and, in the stomach, hydrochloric acid. Surrounding the mucosa is the submucosa, a layer containing blood vessels, lymph vessels, and nerves.
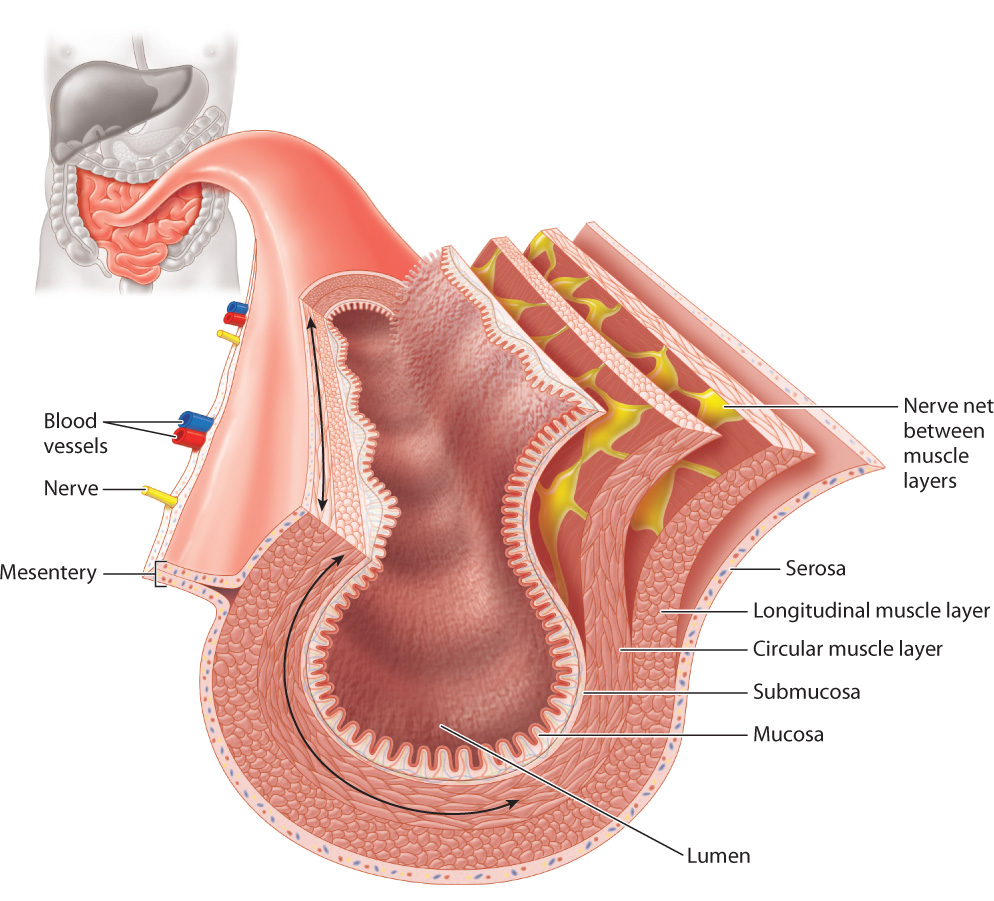
Outside these layers are two smooth muscle layers (Fig. 40.20). An inner circular muscle layer contracts to reduce the size of the lumen. An outer longitudinal muscle layer contracts to shorten small sections of the gut. These two muscle layers contract alternately to mix gut contents, acting as a traveling wave during peristalsis to move the contents along the digestive tract from compartment to compartment. Between the two muscle layers are autonomic nerves that control the contractions of the two sets of smooth muscle.
An outer layer of cells and connective tissue called the serosa covers and protects the gut (Fig. 40.20). The gut is supported in the abdominal cavity by a membrane called the mesentery, through which blood vessels, nerves, and lymph travel to supply the gut.
40.4.7 Plant-eating animals have specialized digestive tracts that reflect their diets.
Herbivorous (that is, plant-eating) animals evolved specialized digestive tracts that enhance their ability to digest cellulose and other plant compounds. Most herbivores, such as cows and termites, lack cellulase, the enzyme that breaks down cellulose. Instead, they have specialized compartments in their digestive tract that contain large bacterial populations, which do produce cellulase. The bacteria are able to digest the cellulose, a main constituent of plants. This association of herbivores and their gut microbes is another example of a symbiotic relationship.
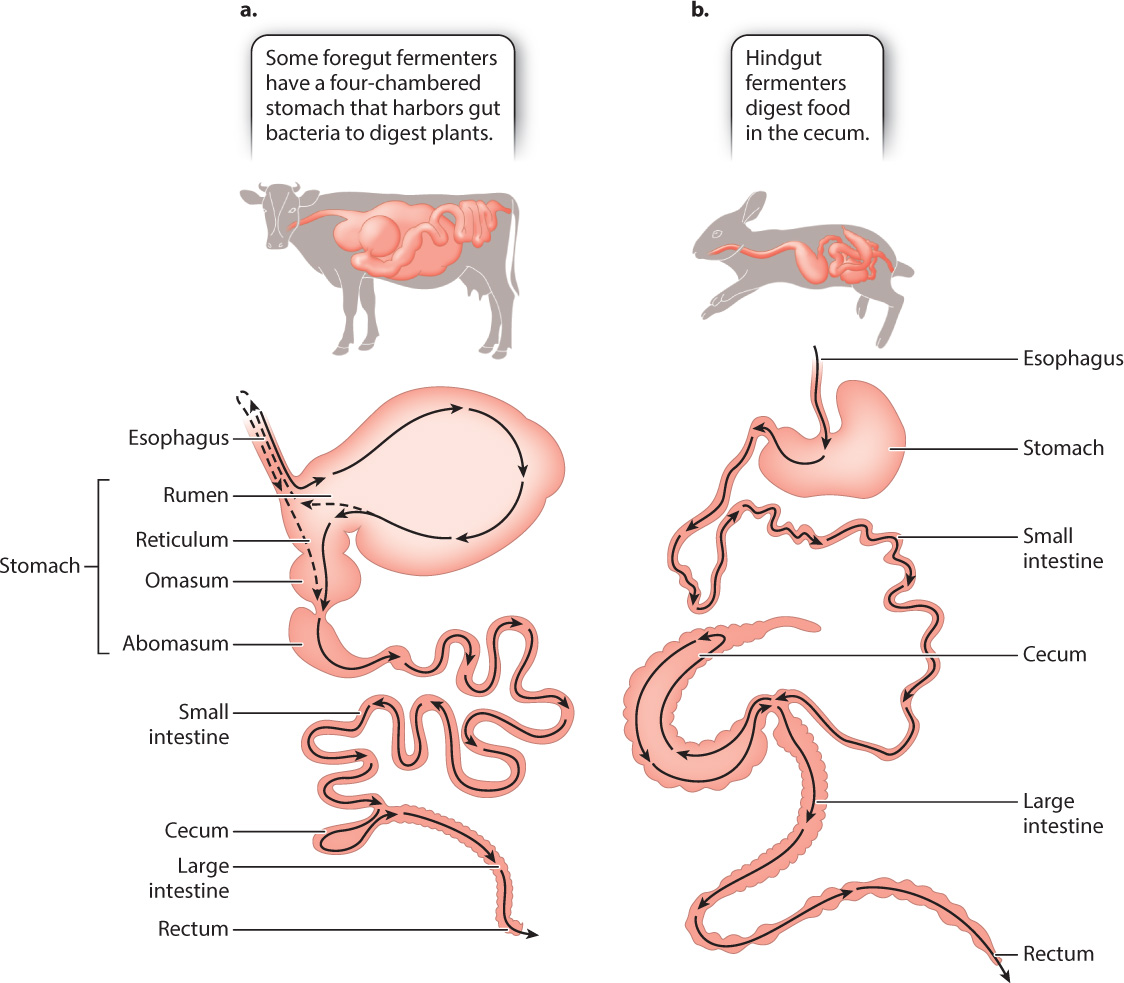
Rather than the single stomach of other mammals, ruminants (cattle, sheep, and goats) have a four-chambered stomach that is highly specialized to enhance the ability of their gut bacteria to digest plants (Fig. 40.21a). The first two chambers—the rumen and the reticulum—harbor large populations of anaerobic bacteria that break down cellulose. To facilitate breakdown, the rumen contents are frequently regurgitated (in a process called rumination) for further chewing in the mouth. This repeated chewing decreases the size of the plant fibers, increasing the surface area available for bacterial digestion in the rumen. The bacteria break down cellulose by fermentation, and some of the products are used as nutrients by the host. (The site of fermentation provides the name “foregut fermenters” for these mammals.) Carbon dioxide and methane gas are also produced as a result of bacterial fermentation. The amount of methane produced by domesticated ruminants is second only to the production of industrial methane. Since methane is a greenhouse gas that traps heat from the sun, methane production by cattle contributes to atmospheric increases in methane and global climate change (Chapter 25).
40-22
After leaving the reticulum, the mixture of food and bacteria passes into a third chamber, the omasum, where water is reabsorbed. It then enters the abomasum, the last of the four chambers, where protein digestion begins as it does in other mammals. The acid and protease secretions kill the bacteria, and the nutritional products are absorbed in the small intestine. The bacteria are an important protein source for the ruminant, yielding as much as 100 g of protein each day for a cow. This is important because plants are a poor source of protein but contain considerable nitrogen that the bacteria use to synthesize their own amino acids. The bacteria lost to digestion are balanced by the continual reproduction of bacteria in the rumen.
In contrast to the foregut fermentation of cows and sheep, other mammalian herbivores, such as koalas, rabbits, and horses, digest the plant material they eat by hindgut fermentation (Fig. 40.21b). Hindgut fermentation occurs in the colon and in the cecum, a chamber that branches off the large intestine. Because the fermentation products released from the cecum of a koala or horse have already passed through the small intestine, the main site of nutrient absorption, hindgut fermentation is less efficient than foregut fermentation in terms of nutrient extraction.
In humans and other animals such as lemurs and squirrels, the cecum is present but small and includes the appendix, a narrow, tubelike structure that extends from the cecum. The appendix is an example of a vestigial structure, a structure that has lost its original function over time and is much reduced in size. Although the appendix has no clear function in nonherbivorous animals, evidence suggests it may play a role in the immune system. The appendix can cause damage to the organism if it becomes infected by partially digested material falling into it. Unless the infected appendix is removed, it can burst, releasing gut contents into the abdominal cavity, a condition that can be life threatening.
40-23
Charles Darwin put forth a hypothesis explaining how the appendix might have evolved to take on its current form in humans. He suggested that in the ancestors of modern humans the cecum once harbored bacteria that aided in the digestion of plant material. However, as their diet changed, they became less reliant on plant material such as cellulose to meet their nutritional needs, so the cecum became smaller and lost its function over time, becoming what we see today as the appendix. Vestigial structures, such as the appendix in humans, remnants of hind limbs in whales, small eyes in cave-dwelling animals, and wings in flightless birds, provide an illuminating window into the past and strong evidence for evolution.