42.3 HUMAN REPRODUCTIVE ANATOMY AND PHYSIOLOGY
In this and the next section, we continue to narrow our focus in order to consider reproduction in more detail. Here, we cover human reproduction and development. While many aspects of human reproduction are shared with other mammals, others are unique. Our focus on human reproduction allows for a detailed treatment of the topic in one organism and also gives us a chance to explore ways we intervene in the process to promote, prevent, and follow pregnancy. Humans, like all mammals, are amniotes. They reproduce sexually, use internal fertilization, and are viviparous.
42.3.1 The male reproductive system is specialized for the production and delivery of sperm.
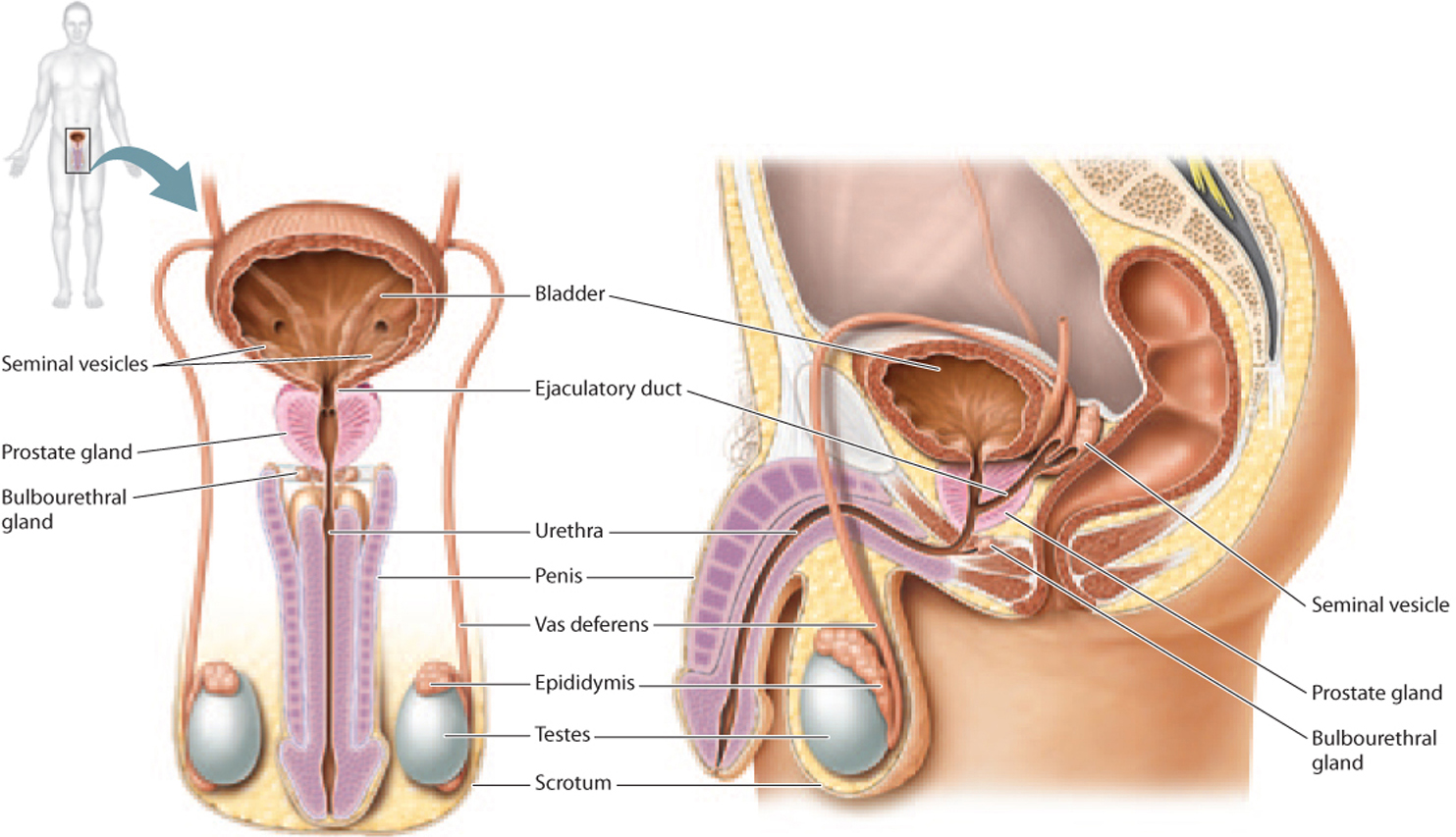
Let’s start by considering the human male reproductive system. Male gametes, called spermatozoa, or sperm, are produced in the two male gonads, or testes (singular, testis; Fig. 42.11). Over the course of his life, a man is capable of making hundreds of billions of sperm. In fact, a single ejaculate typically contains several hundred million sperm. If it takes just one sperm to fertilize an egg, why do males make so many? A likely explanation involves sperm competition. In species like humans, where females can mate with more than one male, a male’s reproductive success is at least partially correlated with the number of sperm—the more sperm, the better chance of fertilizing the egg. A second explanation relates to the female reproductive tract, which is hostile to sperm; many sperm are needed to ensure that at least one makes it to the egg. Of the several hundred million sperm released by a male during a single ejaculation, only several hundred sperm make it to the egg.
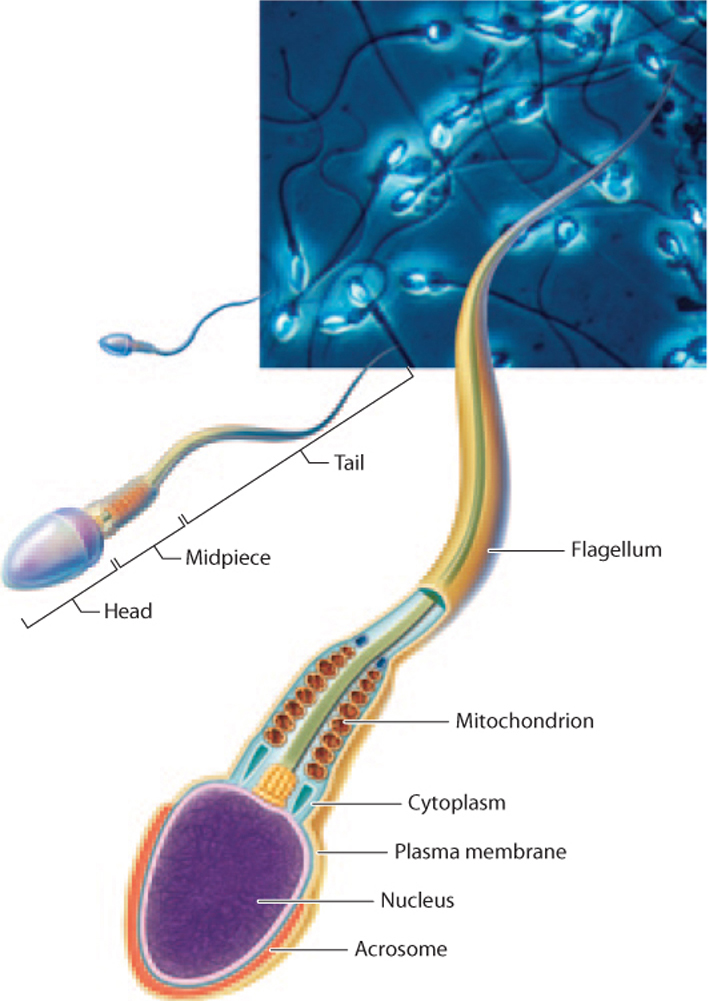
Sperm are haploid cells with a unique shape and properties adapted for their function of delivering genetic material to the egg (Fig. 42.12). They have a small head containing minimal cytoplasm and a very densely packed nucleus. The head is surrounded by a specialized organelle, the acrosome, that contains enzymes that are used by sperm to transverse the outer coating of the egg. Sperm have a long tail, or flagellum, which moves the cell by a whipping motion powered by the sperm’s mitochondria.
The testes are located outside the abdominal cavity in a sac called the scrotum (see Fig. 42.11). This location is critical for the production of sperm, which require cooler temperatures than are found in the abdominal cavity. However, the testes of some mammals, such as elephants, whales, and bats, are located inside the abdominal cavity, indicating that there are species-specific differences in sperm temperature requirements.
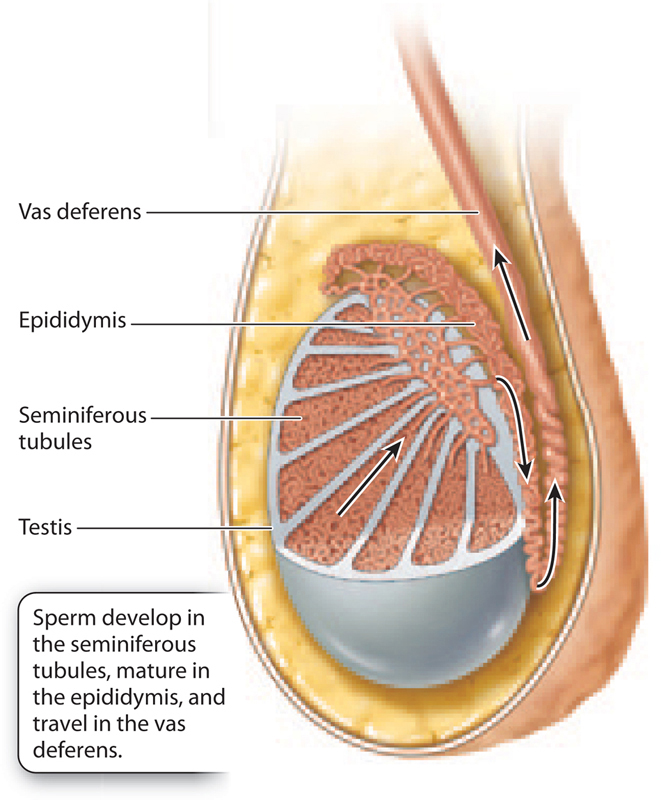
Sperm are produced within the testes in a series of tubes called the seminiferous (“seed-bearing”) tubules (Fig. 42.13). This is where diploid cells undergo meiosis to produce haploid sperm. When sperm are first produced, they are not fully mature: They are unable to move on their own and incapable of fertilizing an egg. They mature as they move through the male reproductive system.
From the seminiferous tubules, sperm travel to the epididymis, where they become motile (Fig. 42.13). This is also where sperm are stored prior to ejaculation. From there, sperm enter the vas deferens, a long, muscular tube that follows a circuitous path, starting off in the scrotum, passing into the abdominal cavity, running along the bladder, and then connecting with the ejaculatory duct (see Fig. 42.11). The two ejaculatory ducts merge at the urethra, just below the bladder. The urethra is used by both the urinary and reproductive systems in males.
Along this path, there are several exocrine glands that produce components of semen, the fluid that nourishes and sustains sperm as they travel in the male and then female reproductive tracts. The first is the prostate gland, located just beneath the bladder (see Fig. 42.11). The urethra and ejaculatory ducts pass directly through this gland. The prostate gland produces a thin, slightly alkaline fluid that helps maintain sperm motility and counteracts the acidity of the female reproductive tract. Two seminal vesicles, located at the junction of the vas deferens and the prostate gland, secrete a protein- and sugar-rich fluid that makes up most of the semen and provides energy for sperm motility. Finally, the bulbourethral glands, below the prostate gland, produce a clear fluid that lubricates the urethra for passage of sperm.
The urethra is enclosed in the penis, the male copulatory organ (see Fig. 42.11). When a male is sexually aroused, the penis becomes erect as a result of changes in blood flow—the arteries become dilated and the veins compressed. During ejaculation, semen is expelled from the penis.
The male reproductive system is well adapted for the production, storage, and delivery of sperm. Therefore, male contraceptive methods focus on blocking sperm. For example, a condom is a sheath of rubber or latex that covers the penis and blocks sperm from entering the female during sexual intercourse.
Question Quick Check 2
eXE14IDpEuMyrZmWe5Ft26REZPGUiKX9h+/7rf/WsU722EP4jwPJ1+o28es0HroO6SAlm+bsKt40hawgEXZQs7uEBWuwEh3WuRUSFJuEkuDMHLt/lanvvkewlYj+xjTlThe human penis is a relatively simple organ, consisting of a shaft and a head called the glans penis. In other animals, the penis can be quite elaborate, with spikes, hooks, and grooves, whose function is to stimulate the female, grip the female, or in some cases even remove sperm from a previous copulation. In marsupial mammals like kangaroos and koalas, males have forked penises, and in snakes and lizards, males have two penises, called hemipenes. In insects, the external genitalia are often distinct enough to be used in species identification. By contrast, most species of birds do not have a penis.
42.3.2 The female reproductive system produces eggs and supports the developing embryo.
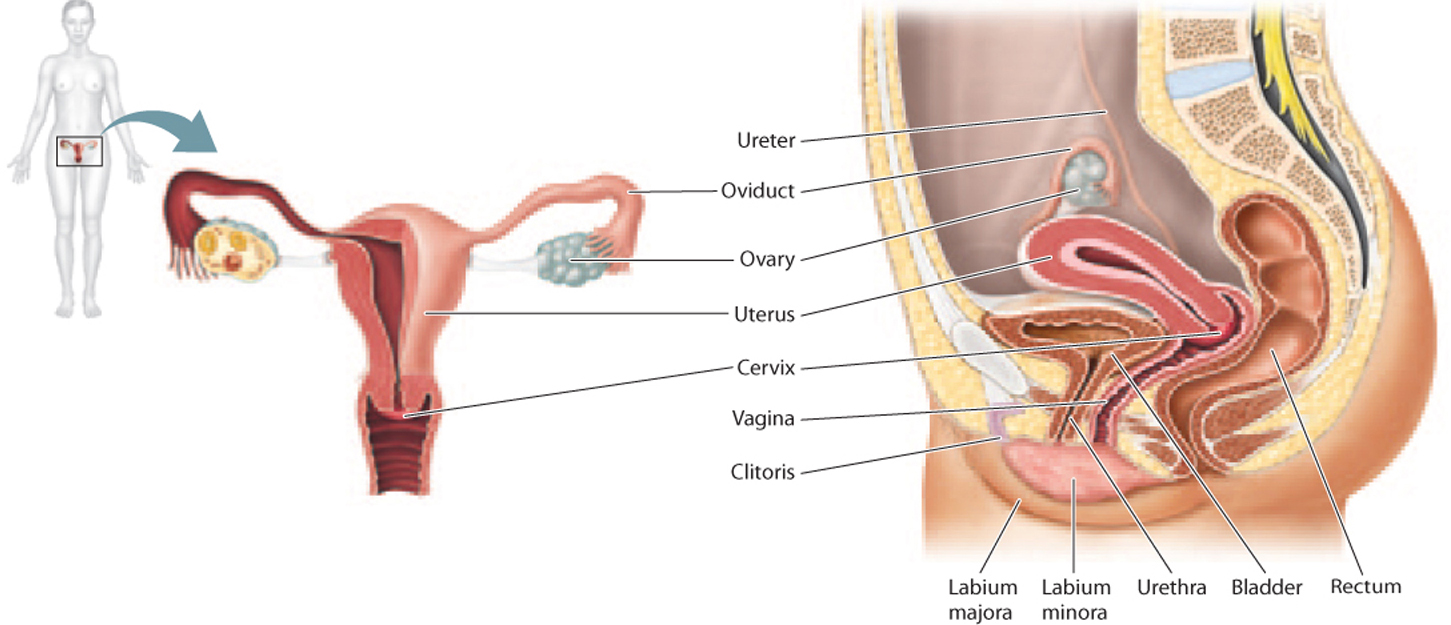
The female reproductive system, like the male reproductive system, produces gametes, but it is also specialized to support the developing embryo. Developing female gametes, called oocytes, which mature into ova, are produced in the two female gonads, called ovaries (Fig. 42.14), and are released monthly in response to hormones (discussed in the next section).
The oocyte is a remarkable and unique cell (Fig. 42.15). In contrast to sperm, which are small and motile, the oocyte is the largest cell by volume in the human body and is visible to the naked eye; it is about the size of the period at the end of this sentence. Its large size reflects the presence of large amounts of cytoplasm containing molecules that direct the early cell divisions following fertilization and food supply in the form of yolk. While a male produces millions of sperm, a female usually produces just one oocyte each month, as she will nourish and support the developing embryo if the oocyte is fertilized.
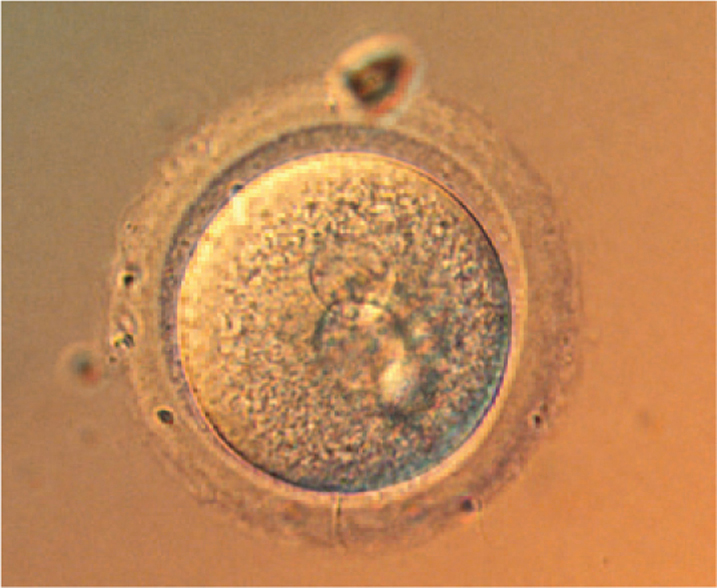
Following release from the ovary, the oocyte travels through the fallopian tube, or oviduct (see Fig. 42.14). There are two fallopian tubes, one on each side. The fallopian tubes have featherlike projections on their ends that help channel the egg into the tube and not into the abdominal cavity.
The oocyte travels from the fallopian tube to the uterus, or womb (see Fig. 42.14). The uterus is a hollow organ with thick, muscular walls that is adapted to support the developing embryo if fertilization occurs and to deliver the baby during birth. In humans, the uterus is usually a single pear-shaped structure, but it can sometimes have two horns, which can interfere with pregnancy. Other mammals differ in the shape of the uterus, ranging from pear-shaped (humans and other primates), to two horns (pigs, horses), to more completely divided but still connected at the base (cats, dogs), to completely separate (rodents, rabbits).
The end of the uterus is called the cervix, or neck, of the uterus (see Fig. 42.14). Under the influence of hormones, the cervix produces different kinds of mucus capable of either blocking or guiding sperm through the cervix.
The uterus is continuous with the vagina, or birth canal (see Fig. 42.14). The vagina is a tubular channel connecting the uterus to the exterior of the body. It is highly elastic, allowing it to stretch during sexual intercourse and birth.
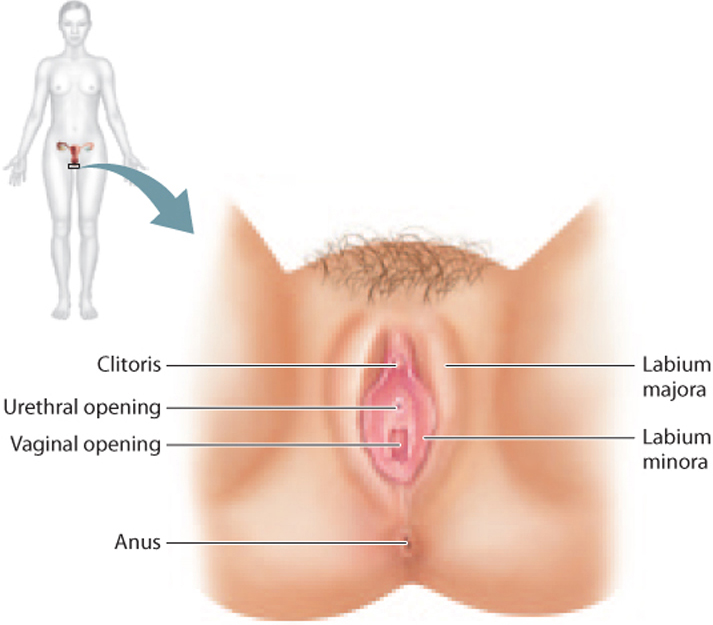
The external genitalia of the female are collectively called the vulva (Fig. 42.16). The vulva includes two folds of skin, the labia majora and labia minora. The labia minora meet at the clitoris, the female homolog of the glans penis.
42.3.3 Hormones regulate the human reproductive system.
The male testes and female ovaries not only are the site of gamete production, but also are part of the endocrine system (Chapter 38). That is, they respond to hormones and in turn secrete hormones. These hormones play key roles during embryonic development and puberty, and also in the adult. Hormonal control of reproduction is critically important, as it allows reproduction to take place only when, for example, a woman is able to support the developing embryo.
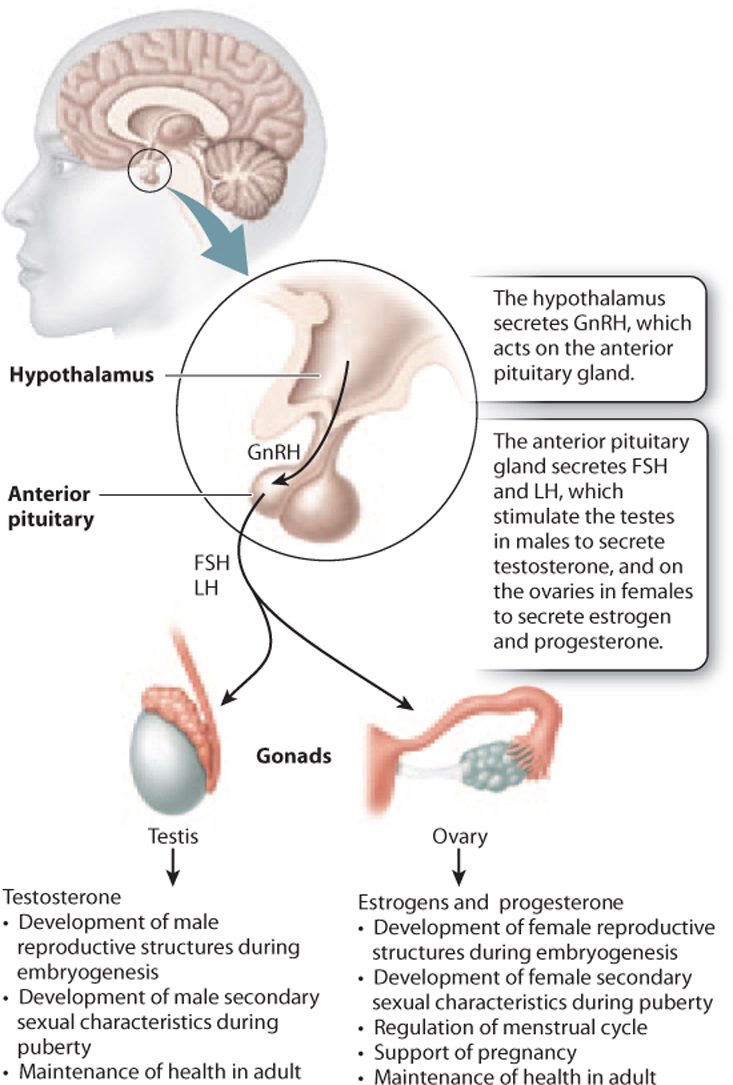
The master regulator of the endocrine system is the hypothalamus, which is located in the brain (Chapter 38). The hypothalamus secretes hormones that act on the pituitary gland, which in turn releases hormones that act on target organs (Fig. 42.17). In the case of the reproductive system, the hypothalamus releases gonadotropin-releasing hormone (GnRH), which stimulates the anterior pituitary gland to secrete luteinizing hormone (LH) and follicle-stimulating hormone (FSH). These hormones then act on the male and female gonads. In males, these hormones are secreted almost continually, whereas in females, they are secreted cyclically. In response, the testes secrete the hormone testosterone and the ovaries secrete the hormones estrogen and progesterone.
In males, LH acts on cells in the testes called Leydig cells, which secrete testosterone. Testosterone is a steroid hormone derived from cholesterol that plays key roles in male growth, development, and reproduction. Testosterone is important during embryogenesis to direct development of the male reproductive organs. At puberty, levels of testosterone increase, leading to the development of male secondary sexual characteristics, traits that characterize and differentiate the two sexes but that do not relate directly to reproduction. For males, these include a deep voice, facial, body, and pubic hair, and increased muscle mass. Testosterone together with FSH act on another population of cells in the testes called Sertoli cells to stimulate sperm production. Sertoli cells, located in the seminiferous tubules, support the process of sperm production. In this way, hormones help to regulate the timing of sexual maturity.
In females, FSH acts on cells surrounding the oocyte called follicle cells, which support the developing oocyte and secrete estrogen. At puberty, an increase in the level of estrogen leads to the development of female secondary sexual characteristics, such as enlargement of the breasts, growth of body and pubic hair, and changes in the distribution of muscle and fat. Also at puberty, females begin a monthly cycle, the menstrual cycle, in which oocytes mature and are released from the ovary under the influence of hormones (Fig. 42.18). The menstrual cycle consists of cyclical and coordinated changes in the ovaries and uterus, timing the release of an oocyte from the ovary with the growth of the uterine lining so it can support the developing embryo if fertilization occurs.
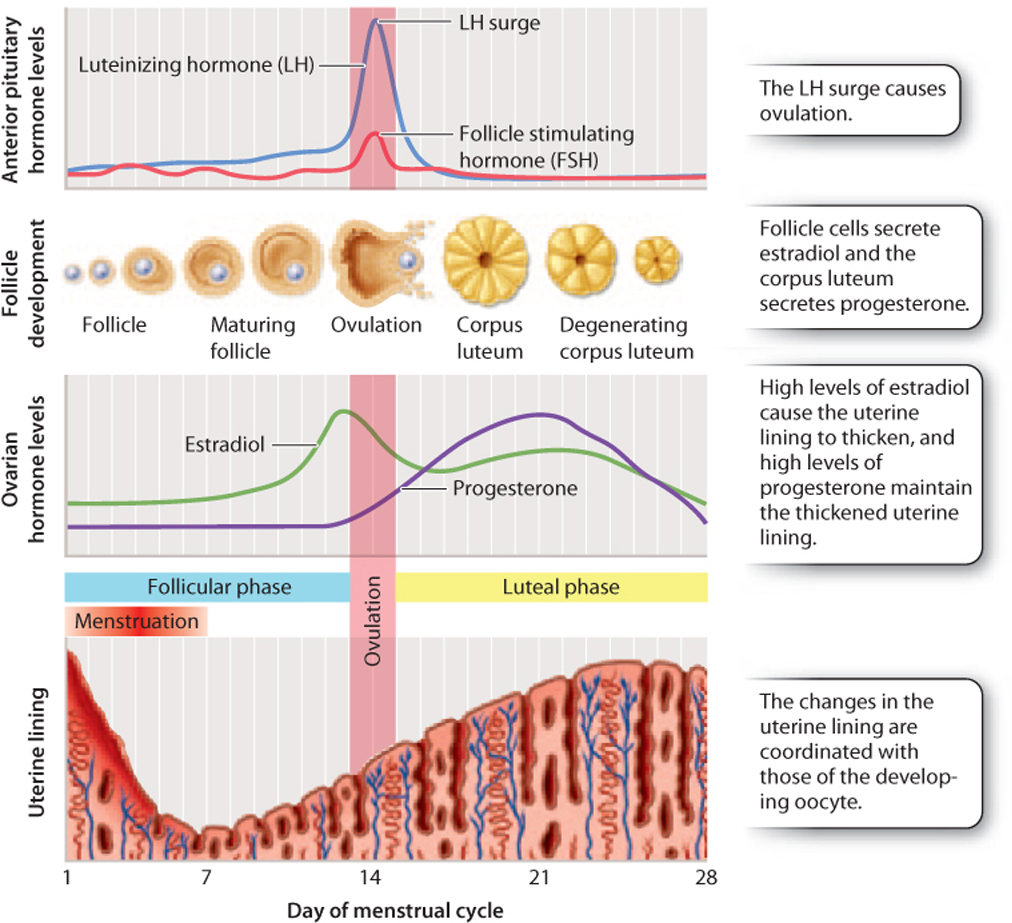
The menstrual cycle has two phases, the follicular phase and the luteal phase. In the follicular phase, FSH acts on a subset of follicle cells called granulosa cells. As a result, several oocytes begin to mature, but usually only one of these becomes completely mature and the others die off. The granulosa cells secrete a form of estrogen known as estradiol. Estradiol in turn acts on the lining of the uterus, causing it to thicken.
As a result in part of rising estradiol levels, there is a rapid increase followed by a sharp decrease in the level of LH produced by the anterior pituitary gland. This surge causes ovulation, the release of the oocyte from the ovary. Ovulation marks the beginning of the luteal phase. The follicle cells, now devoid of the oocyte, are converted to a structure known as the corpus luteum, literally “yellow body,” so called because of its appearance. The corpus luteum is a temporary endocrine structure that secretes the hormone progesterone, which maintains the thickened and vascularized uterine lining.
The oocyte, meanwhile, is swept into the fallopian tube and travels to the uterus. If it is fertilized, the developing embryo implants into the uterine lining. The corpus luteum continues to secrete progesterone, and is first maintained by LH and then by the hormone human chorionic gonadotropin (hCG), which is released by the developing embryo. Eventually, the placenta takes over estrogen and progesterone production to maintain the uterine lining and stimulate growth of the uterus. High levels of estrogen and progesterone during pregnancy also block ovulation because they suppress the release of GnRH, FSH, and LH.
If the oocyte is not fertilized, the corpus luteum degenerates, estrogen and progesterone levels drop, and the uterine lining is shed. The monthly shedding of the uterine lining is known as menstruation. Menstrual cycles usually start occurring around age 12 in the United States and continue until approximately age 45 to 55. The cessation of menstrual cycles, called menopause, results from decreasing production of estradiol and progesterone by the ovaries.
As we have seen, the brain, anterior pituitary, ovaries, uterus, and placenta pass hormonal messages back and forth to make sure that behavior, gamete release, the uterine lining, and embryo development are coordinated. If hormone levels are low or absent, as occurs normally before puberty or as the result of malnutrition or some diseases, ovulation does not occur. Similarly, too much hormone at the wrong time can interfere with fertility, which is the basis for oral contraceptive pills (“the pill”). Oral contraceptive pills contain various combinations of synthetic estrogens or progesterones or both.
Question Quick Check 3
dChxb0cpeJ0MNQKxHgc0+XozO7Sx0hGX623kZ30mEXxq90xfWeGrRWerFODZZi+WldaFlxLu7RpfMvyWwSQ4OXgGTyvECSDrsdWAOaV9BZGv8H4DxfU7f0Zl11UPWGezaySihknkFWYoTzzBHumans and chimpanzees both have menstrual cycles. Other placental mammals have an estrus cycle, which is characterized by phases in which females are sexually receptive. In these mammals, the uterine lining is reabsorbed instead of shed if fertilization does not occur.