45.3 LEARNING
Genes are not the only determinants of behavior. We all know that experience often leads to changes in behavior. We call this process learning. Humans are extraordinary learners, but even many relatively simple organisms respond to experience. Researchers have identified several types of learning. The categories are not strict, and there is some overlap among them, but they are a useful way to organize our thinking and give us deeper insights into the subject.
45.3.1 Non-associative learning occurs without linking two events.
Non-associative learning is learning that occurs in the absence of any particular outcome, such as a reward or punishment. One type of non-associative learning is habituation, which is the reduction or elimination of a behavioral response to a repeatedly presented stimulus. Chicks presented with silhouettes flying overhead provide an example. Initially, any overhead silhouette provokes a defensive, crouching posture, but eventually, chicks habituate to overhead silhouettes that have proved not to be threatening. In the absence of any consequences, chicks no longer crouch in response to a harmless silhouette passing repeatedly overhead.
Sensitization is another form of non-associative learning. Sensitization is the enhancement of a response to a stimulus that is achieved by presenting a strong or novel stimulus first. This pre-stimulus makes the animal more alert and responsive to the next stimulus. The sea slug, Aplysia, a model organism among scientists interested in the neuronal basis of behavior, exhibits sensitization. Aplysia withdraws its gills in response to a touch on the siphon, the tube through which it draws water over its gills. Interestingly, if the animal is given a weak electric shock first, its response to a touch on its siphon is much more rapid, suggesting that the shock has made the slug more sensitive to later stimuli.
45.3.2 Associative learning occurs when two events are linked.
Associative learning (also called conditioning) occurs when an animal learns to link (or associate) two events. Perhaps the most famous example of conditioning is Ivan Pavlov’s dogs. Pavlov first presented the dogs with meat powder, and they salivated in response. He then presented the dogs with an additional cue, a ringing bell, whenever he presented the dogs with meat powder. After repeated experience of the two stimuli together, the dogs salivated at the sound of the bell alone, in expectation of the meat reward. This form of conditioning, in which two stimuli are paired, is called classical conditioning. In this case, a stimulus that leads to a behavior (the meat powder) was paired with a neutral stimulus (the bell) that initially had nothing to do with salivation. Eventually, a novel association is made between the sound of the bell and food, and the sound of the bell alone elicits salivation.
A second form of associative learning involves linking a behavior with a reward or punishment. If the behavior is rewarded (positively reinforced), it is more likely to occur the next time around. It is not just out of the goodness of her heart that the trainer at Sea World gives a sea lion or dolphin a fish reward at the end of each trick that the animal performs. However, if the behavior is punished (negatively reinforced), the response becomes less likely. This form of associative learning is called operant conditioning.
In classical conditioning, an association is made between a stimulus and a behavior, whereas in operant conditioning, an association is made between a behavior and a response. In addition, operant conditioning involves a novel, undirected behavior that becomes more or less likely over time. Consider a rat in a cage with some levers in it, each a different color. The rat has no impulse to press any of the levers, but by chance, it presses the red one. A food reward tumbles into the cage. Eventually the rat learns to associate pressing the red lever with the food reward; it has been operantly conditioned. It is thought that play in young animals provides them with the opportunity to explore their environment much in the same way as the rat does when first placed in the cage. Like the rat, they may find that some behaviors are more rewarding or more punishing than others.
Question Quick Check 2
nNMm0Lt2aHOV2NEXMT72UK2QSLNJ9t/S05fgG1fshCYAcVqLh0apouY9KrWZdAvEKrs6i3UlIUzFdRpuNkvp5QM4svdmp3ZcZsqlFol6g4yscTuF60aa4OFH6Wz6blr4mRXfjbhgdjk/rKq4B5kGFAxbxwyZyhFuvUozGXaMEu+1w6tbY/Umryu0iHJJpoXMbXm9PMfgmic=45.3.3 Learning takes many forms.
The capacity to learn often has adaptive functions. Much learning, including human learning, is based on imitation: One individual copies another. Learning by imitation was famously observed in the days when people in Britain had milk deliveries left outside on the doorstep: A number of birds, including Great Tits and Blue Tits, learned to open foil-topped milk bottles, and this behavior spread rapidly through populations of these birds as naïve birds copied the successful birds’ approach. Individuals can also learn by imitating individuals of a different species. An octopus can learn to open a jar with a reward inside by watching a human or another octopus do it.
The capacity to learn a particular task seems in many instances to be innate. Female digger wasps are foragers. Tinbergen showed that a digger wasp learns the landmarks around her nest and then uses this information to find her way back to it from hunting. In his experiment, illustrated in Fig. 45.10, Tinbergen placed a ring of pine cones around a wasp’s nest and, after she had flown out and returned a few times, moved the circle of pine cones to the side of the nest. The wasp flew past her own nest and went directly to the center of the ring. Such innate responses improve the survival or reproductive success of the individual.
FIG. 45.10: To what extent are insects capable of learning?
BACKGROUND European digger wasps, Philanthus triangulum, live in the sand. These wasps are sometimes called “bee wolves” because they specialize on hunting for honeybees to feed their developing young. After mating, each female digs a long burrow with a few chambers at the end where she lays her eggs. She then forages for honeybees that she brings back to these chambers for her larvae to eat. The wasp faces a navigational challenge: Having captured her prey, how can she find her way back to and recognize her nest? Niko Tinbergen noticed that wasps lingered briefly in the vicinity of a new nest before heading off to hunt, and thought that they were learning local landmarks associated with the nest.
HYPOTHESIS The wasp learns visual cues around her nest to help her locate it upon her return.
METHOD Tinbergen recognized that a good test of the learning abilities of an insect should take place in its natural environment. He combined his skills as a naturalist and as an experimentalist to devise an elegant demonstration of the way in which female wasps learn landmarks for navigation. Tinbergen’s approach was simple. He placed a ring of pine cones around the nest of a wasp, and then, once she had left to hunt, he shifted them to a new location away from the nest entrance. If visual cues are key, the wasp would return to the displaced pine cone ring. On the other hand, if cues are, for example, olfactory, she would return directly to the nest.
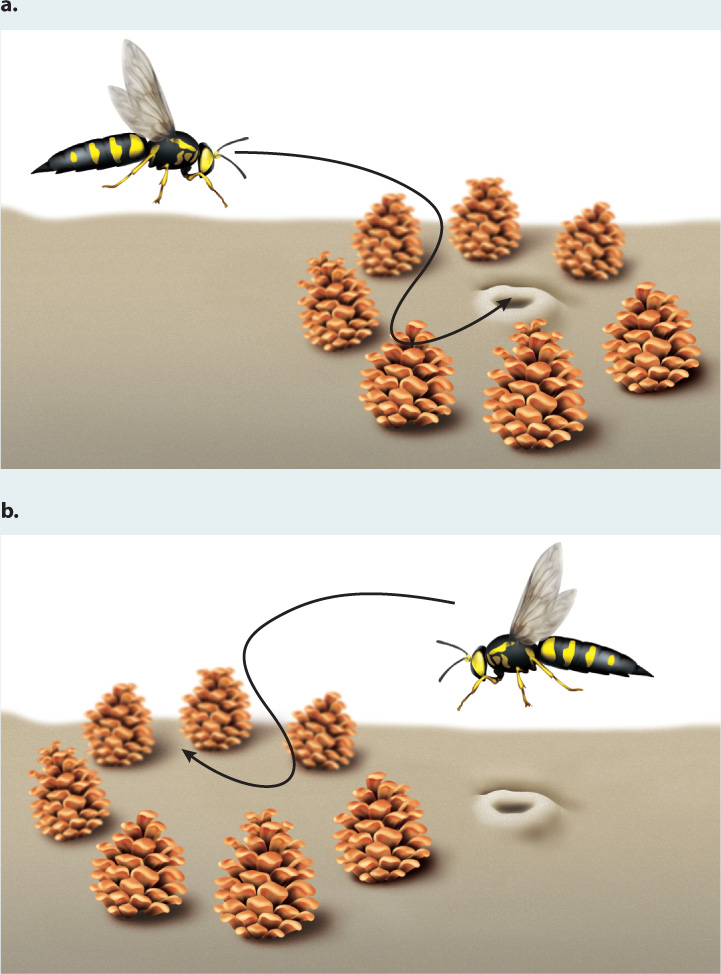
RESULTS Females carried out a brief landmark-learning flight on departure from the nest (Fig. 45.10a). When the landmarks were displaced, the females returned to the wrong location (Fig. 45.10b).
CONCLUSION Female digger wasps learn and then use local landmarks, such as a ring of pine cones, as cues to the nest location.
SOURCE Tinbergen, N. 1958. Curious Naturalists. New York: Basic Books.
This adaptive aspect of animal learning is also revealed by taste aversion experiments, in which an animal typically learns to avoid certain flavors associated with a negative outcome. Rats learn to avoid flavored water if consuming it is associated with nausea. However, they do not learn to avoid flavored water if it is associated with a different kind of negative reinforcement, such as a mild electric shock. Rats, it turns out, can make some associations, but not others. The ones they can learn are the biologically meaningful ones, those that favor survival. In the course of evolution, the rat’s ancestors encountered poisoned food that resulted in nausea and the aversion response evolved. Until humans started doing experiments on them, rats had never encountered a bad meal that resulted in an electric shock. It is not surprising, then, that the ability to pair these two phenomena never evolved. These experiments also show that the specific evolutionary history of the species in question matters when analyzing animal behavior.

In addition to adaptive predispositions for what can be learned and not learned, many species exhibit predispositions for when learning takes place. This is particularly evident in imprinting, a form of learning typically seen in young animals in which they acquire a certain behavior in response to key experiences during a critical period of development. Konrad Lorenz made imprinting famous by exploiting the observation that newly hatched goslings and ducklings rapidly learn to treat any animal they happen to see shortly after hatching as their mother. Lorenz found that, if he was the first person the hatchlings saw, they would follow him as though he were their mother (Fig. 45.11). This behavior is adaptive because the first being that a hatchling normally sees is a parent. This type of imprinting is called filial imprinting, and it is most common in species whose offspring leave the nest and walk around while still young, like chicks and ducklings. Filial imprinting is rare in species of birds whose young stay in the nest until they are able to fly away.
Experiments have shown that filial imprinting typically occurs during a specific, sensitive period in the animal’s life and that the results are usually irreversible. After Lorenz’s baby ducks had imprinted on him, they would not change their minds about who their parent was even when presented with their real mother duck. The timing of the sensitive period varies from species to species. In some cliff-nesting sea birds, it has been shown that imprinting on auditory stimuli (like the call of the parents) begins while the chick is still in the egg.