45.4 ORIENTATION, NAVIGATION, AND BIOLOGICAL CLOCKS
Learning is a form of information processing, in which experience shapes behavior. At a mechanistic level, it typically involves a change in the strength of connections between neurons. The central nervous system of even relatively simple animals is capable of extraordinary feats of information processing. The ways in which animal nervous and endocrine systems and the input of experience are integrated to generate adaptive behaviors are remarkable. Some of these adaptive behaviors include ways of moving, navigating, and keeping time. Here, we look at how organisms integrate environmental stimuli to produce these behaviors.
45.4.1 Orientation involves a directed response to a stimulus.
Even the simplest bacteria and protozoa are capable of moving in response to stimuli. A Paramecium that finds itself in an unfavorable environment, such as water that is too warm or too salty, increases its speed and begins to make random turns. When it finds favorable conditions, such as cooler water, it slows and reduces its turning rate. These random, undirected movements are termed kineses.
In contrast, taxes (singular, taxis) are movements in a specific direction in response to a stimulus. An interesting example of a taxis is movement oriented to a magnetic field, a behavior called magnetotaxis—the term was first used by the American microbiologist Richard Blakemore in 1975. Blakemore had found that anaerobic bacteria in the genus Aquaspirillum, which swim by means of flagella, tend to swim toward magnetic north. These bacteria can be attracted to the side of a dish with a bar magnet. Little bits of magnetized iron oxide, arranged in a row inside the bacterial cell, allow the bacteria to sense the magnetic field. Blakemore hypothesized that the bacteria swim north in order to swim deeper. In the northern hemisphere, the north magnetic pole is inclined downward. At Woods Hole, Massachusetts, where these bacteria were found, the magnetic pole is at about a 70-degree incline. Since the bacteria are anaerobic, they must stay buried in sediment, and they remain buried by moving downward along the magnetic gradient.
To test his hypothesis, Blakemore looked for and found bacteria in New Zealand that exhibited similar behavior to the Woods Hole bacteria except that they swam toward the south magnetic pole, bringing them downward in the southern hemisphere. Finally, he took New Zealand cultures back to Woods Hole, where the bacteria swam up into the oxygen and died. These experiments demonstrated that bacteria are able to sense a magnetic field and move in a directed fashion relative to that field.
45.4.2 Navigation is illustrated by the remarkable ability of homing in birds.
Many animals use environmental cues to migrate long distances. The navigational achievements of homing pigeons are legendary. They can home—that is, find their way back to the place where they are housed and fed—over extremely long distances, even more than a thousand miles. It seems that they use a wealth of cues when homing. A compass may tell you which way is north, but you must also have map information—you must know where you are with respect to your goal—if the compass is to be useful for finding a particular location. So pigeons must have both compass and map senses. That map sense is presumably based on landmarks. Given the long distances the pigeons travel, these landmarks must vary in their type and probably include olfactory as well as visual cues.
The pigeon compass relies on different kinds of information. For example, pigeons can navigate during the day using the sun as a compass, and at night they use the stars. And, like Aquaspirillum, they can detect the Earth’s magnetic field. The magnetic navigation system is presumably important to pigeons on cloudy days. Researchers have performed experiments with pigeons wearing magnetic helmets, which disrupt the pigeons’ ability to detect the Earth’s magnetic field. On cloudy days, the pigeons wearing the helmets are unable to home. On sunny days, however, the pigeons’ sun compass overrides the erroneous magnetic information of the helmets.
The sun compass requires information about time as well. Every hour, the sun moves 15 degrees through the sky. To determine where north is, a pigeon needs information from the sun as well as some way to keep time. It turns out that pigeons have a clock—a biological one.
45.4.3 Biological clocks provide important time cues for many behaviors.
Like us, other animals live in space and time. So far, we have focused on their behavior in space. For many species, time is a life-and-death matter. When to migrate or mate is a critical decision in a seasonal environment. Researchers are beginning to unravel the neural and genetic underpinnings of the clocks in some species. In model organisms like Drosophila, for example, researchers have identified a number of genes that, when mutated, cause the clock to run slow or fast.
A biological clock is a set of molecular mechanisms that cycles on its own and therefore keeps a regular rhythm. It helps to control physiological or behavioral aspects of an animal. Different clocks work on different timescales. Daily cycles are governed by a circadian clock. Circadian clocks regulate many daily rhythms in animals, such as feeding, sleeping, hormone production, and core body temperature. Some species, especially seacoast species living in habitats where the tides are important, time activities by a lunar (moon-based) clock. There are also annual (yearly) clocks. For example, periodical cicadas are insects in the genus Magicicada that have a generation time of either 13 or 17 years: In a given location, there will be a cicada outbreak every 13 or 17 years.
Circadian clocks are observed in many organisms, including plants, fungi, and animals. Humans have their own circadian clocks, as jetlag never fails to remind us. The circadian clock is based on a set of clock genes. The protein products of these genes oscillate through a series of feedback loops in a roughly 24-hour cycle. Thus, when animals that, like us, are active during the day and inactive at night are placed in artificial conditions that are always lit, they continue to follow a basic day–night, active–inactive cycle. The clocks are not perfect, though: As the period spent in constant light is prolonged, the circadian clock drifts slowly until the animal is eventually no longer synchronized with the true day–night cycle.
Biological clocks remain synchronized with the day–night cycle because they are often reset, or entrained, by external inputs. For the circadian clock, light is the primary input, so the natural light–dark cycle keeps the clock from drifting. For clocks related to the seasons, day length (known as photoperiod) is the critical input because it is a good indicator of the time of the year (Chapter 30). For example, many mammals produce their offspring in the spring so that they can grow over the summer, when resources are most abundant. These mammals have been selected over many generations to synchronize their reproduction at a time when the young have the greatest chance of survival. Photoperiod determines the timing of many kinds of behaviors, including migration, development, and reproduction.
The importance of the sun compass coupled with the biological clock in homing pigeons can be demonstrated by experimentally disturbing the birds’ clock, as discussed in Fig. 45.12.
FIG. 45.12: Does a biological clock play a role in birds’ ability to orient?
BACKGROUND One suggestion for how pigeons home is that they use a sun compass. If you are in the northern hemisphere and you know the time is 12 noon, then the sun is due south of you. Orienting yourself by this method is possible only if you know the time, so the question arises whether homing birds have the ability to tell the time. One way to answer this question is to “clockshift” the birds. Researchers raise birds in an artificial day–night cycle that is out of sync with the actual one. When released into a sunlit environment, these birds’ sense of time is shifted by a set number of hours:

HYPOTHESIS If a bird’s ability to home is dependent on an internal clock, clockshifting should affect the bird’s homing ability in a predictable way. Given that the sun travels 360 degrees in 24 hours, a 6-hour clockshift will result in a 90-degree error in homing direction because 360/(24/6) = 90.
EXPERIMENT Birds were clockshifted by raising them in a chamber under an artificial light. Birds from a home loft in Ithaca, New York, were released on a sunny day at Marathon, New York, about 30 km east of Ithaca. Release on a sunny day made it possible for the birds to use the sun’s position to navigate.
RESULTS As expected, the control birds (those that were not clockshifted) were usually good at picking the direction of their home loft, heading approximately westward toward Ithaca. The results for the clockshifted birds were very different: They miscalculated the appropriate direction. These birds headed approximately northward, as shown by the positions of the red triangles on the compass in the figure.
INTERPRETATION Assume the birds are released at 12 noon, when the sun is due south. The control birds know to fly in a direction 90 degrees clockwise from the direction of the sun, but the clockshifted birds “think” it is 6 p.m., so they expect the sun to be in the west. Their 90-degree clockwise correction, then, has them flying due north.
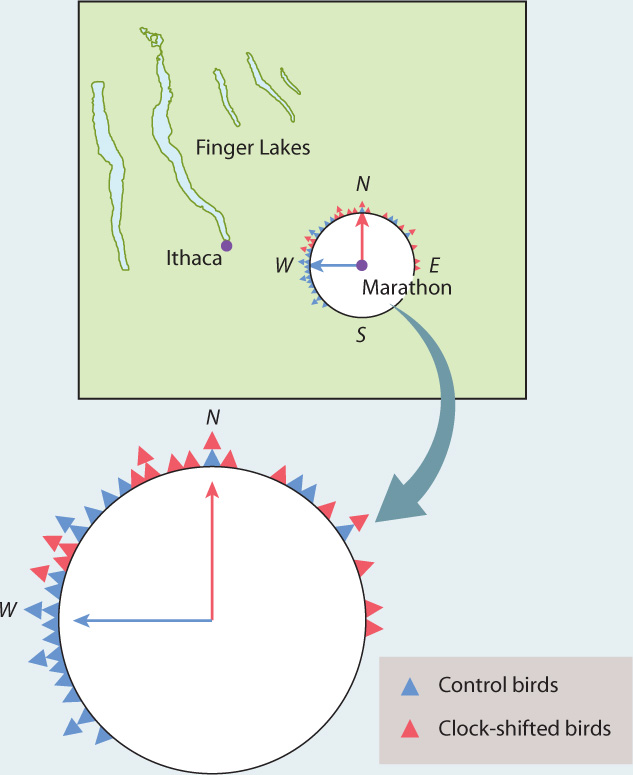
CONCLUSION The clear difference between control and clockshifted birds in the experiment shows that an internal time-based sun compass is an important component of the birds’ homing abilities. However, the scatter of points (for both experiment and control) suggests that other factors are also important. This conclusion is reinforced by the observation that birds home well on cloudy days, when they cannot use a sun compass, suggesting that birds use multiple cues and navigational systems when they are homing.
SOURCE Keeton, W. T. 1969. “Orientation by Pigeons: Is the Sun Necessary?” Science 165 : 922–928