45.5 COMMUNICATION
Up to this point, we have focused on individual behaviors. Behaviors often depend on environmental cues, as we saw with the examples of navigation and keeping time. In addition, behaviors are often shaped by interactions with other individuals. Learning sometimes depends on communication between a teacher and student, and, as anyone knows who has ever watched a mother duck tending her ducklings, communication is central to filial imprinting.
45.5.1 Communication is the transfer of information between a sender and receiver.
The sophistication of animal communication varies enormously, as can be seen by the variety of types of signal illustrated in Fig. 45.13. Even our closest relatives, the chimpanzees, cannot rival the human facility with language, but a number of species have evolved forms of communication that convey remarkably complex and specific information. For example, a vervet monkey that perceives a threat to its group will utter an alarm call that not only warns of a threat, but actually specifies the nature of the threat—whether it is a hawk, leopard, snake, or some other type of predator.

The simplest definition of communication is the transfer of information between two individuals, the sender and the receiver. The sender supplies a signal that elicits a response from the receiver. For example, the bright petals of a flower signal to an insect that nectar and pollen are available. This definition, however, has its problems. An owl hears the rustling of a mouse and responds accordingly. Most people would agree that the owl is not really communicating with the mouse (nor the mouse with the owl). For this reason, some biologists prefer to define communication as attempts by the sender to manipulate in some way the behavior of the receiver.
How has communication evolved? It is thought that communication has often evolved through co-opting and modifying behaviors used in another context. This process is called ritualization, and it involves (1) increasing the conspicuousness of the behavior; (2) reducing the amount of variation in the behavior so that it can be immediately recognized; and (3) increasing its separation from the original function. The scent markings with which mammals mark their territory provide an example. The original function was simply the elimination of waste, but this function has been modified through evolution: Strategically placed marks communicate with other individuals, indicating, for example, the extent of an individual’s territory. The communication advantage is that an intruder detecting a territorial scent may be less likely to invade, thereby avoiding what could be an expensive fight.
Forms of communication have frequently evolved that prevent animals from coming to harm in what has sometimes been called limited-war strategies. Thus, two males, instead of battling it out, may engage in elaborate displays to size each other up, either literally by standing side by side or through displays of physical prowess such as roaring, in order to determine who is dominant. Fighting would be disadvantageous to both since even the winner might be seriously injured. In this case, communication may have evolved as a way for individuals to assess each other.
In some cases, communication in the natural world can be deceitful. A male may attempt to convince other males (or a female) that he is bigger (and stronger) than he really is. Possibly one reason that dogs circling in a fight raise the hair along their back is to inflate their apparent size. A potential prey may attempt to convince a predator that it is not, in fact, prey. This form of deceit is especially evident in some species of butterfly that mimic leaves or bark when their wings are at rest. Alternatively, predators may emit deceitful signals to entice their prey. Females of one species of firefly, for instance, mimic the flashes of the females of another species in order to lure males of the second species, which they then eat.
45.5.2 Some forms of communication are complex and learned during a sensitive period.
Bird song is one of the best known and richest forms of animal communication. Because of the clear connection between sensory perception and motor output involved in this form of communication, songbirds have served as a model system in the study of learning and communication. These songs are complex sequences of sounds, often repeated over and over. Like cricket and frog calls, bird songs are advertisement displays, behaviors by which individuals draw attention to their status (for example, that they are sexually available or are holding territory). Bird songs are typically produced in the breeding season and usually only by males, although in some species females also sing and perform “duets” with their mates.
In many species of birds that sing, some or all of the song is learned, often during a specific, sensitive period. Detailed studies pioneered by British-American neurobiologist and ethologist Peter Marler of the White-Crowned Sparrow, shown in Fig. 45.14, have yielded the following general picture of the process.
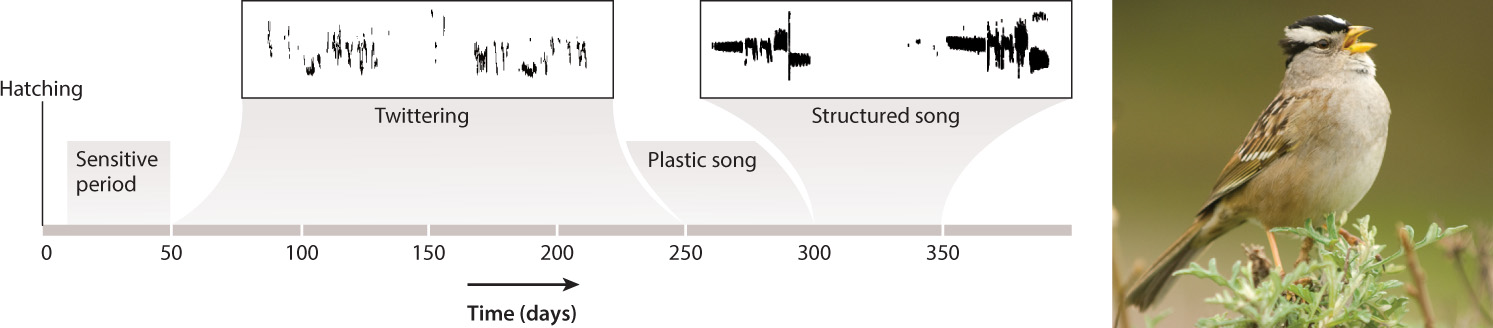
Song in White-Crowned Sparrows is learned by imprinting: The young male hears adult song during a sensitive period, 10–50 days after hatching. Then, shortly afterward, young male White-Crowned Sparrows produce unstructured twittering sounds, known as a subsong, comparable to the babbling of human babies. If deprived of hearing adult song, for example by isolation during the sensitive period, the bird sings for the rest of his life a song not much different from unstructured twittering, even if he hears his father sing both before and after isolation.
Between about 250 and 300 days, the male sings an imperfect copy of his father’s song (known as plastic song), and by about 300 to 350 days after hatching, song acquisition is complete (this song is known as structured song). At this point, even if the bird is deafened in the lab, he will still sing correctly. The song he produces is a precise copy of the one he learned, typically by hearing his father’s song during the sensitive period, complete with most of its individual as well as species-specific characteristics.
So, White-Crowned Sparrows have a programmed predisposition for when song learning takes place. Even more interestingly, what can be learned is similarly constrained. If a tape of another species’ song is played during the sensitive period, even the song of closely related Swamp Sparrows, the White-Crowned male cannot learn that song; his adult song ends up being not much different from his unstructured twittering. However, if he is provided with a live tutor, such as a live male Swamp Sparrow rather than a tape, his ability to learn the other bird’s songs is greatly improved. Furthermore, if he hears tapes of both the song of his own species and that of closely related species played together, he can pick out the correct elements and sing a perfect White-Crowned Sparrow song. Thus, birds of this species preferentially learn their species-specific song, but they cannot sing a normal song without learning it.
45.5.3 Other forms of communication convey specific information.
Honeybees have an elaborate means of communication that is quite different from the sound-based communication of birds. To collect food for a honeybee hive, about 10,000 foragers leave the nest each day, making several million foraging trips in the course of a summer, each of which may take them up to 10 km from the nest. The hive can be considered a kind of giant organism, fixed in one spot but sending out foragers in all directions. As social insect expert and evolutionary biologist E. O. Wilson points out, if honeybee workers were as large as people, they would cover an area the size of Texas in their daily search for food. Given the vastness of this task, it is not surprising that returning workers use a variety of cues to inform outgoing foragers about the location and nature of the food resources they have found.
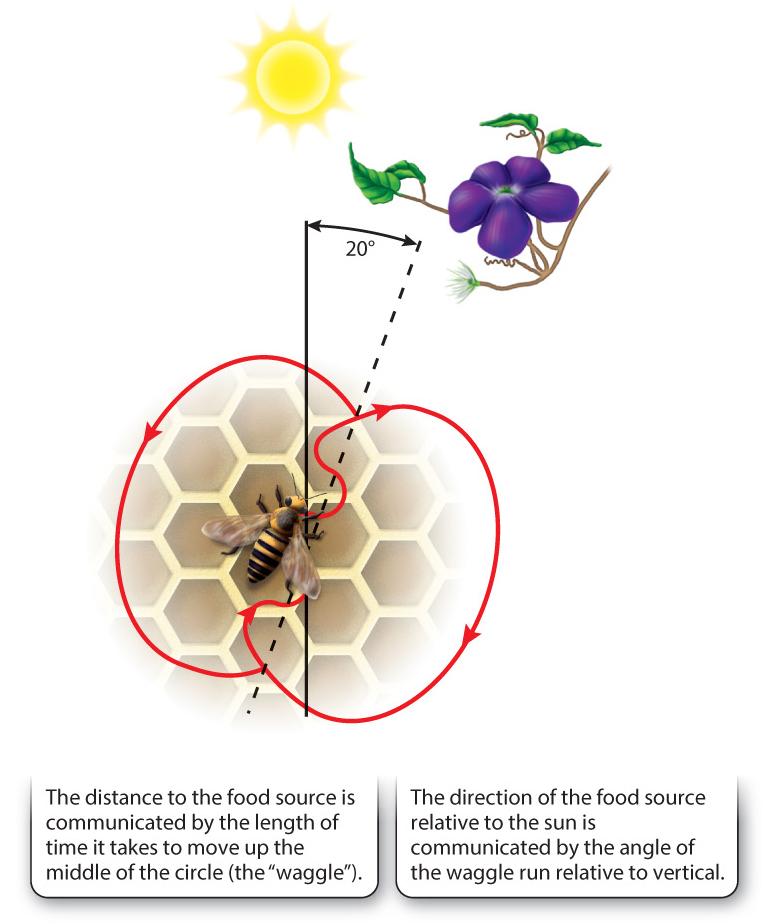
Returning workers pass samples of the collected food to other workers that gather around the forager when she returns to the hive. The returning foragers then begin a series of movements in specific patterns that encode information about the direction and distance of food sources. This is the dance language of honeybees, originally discovered by Karl von Frisch.
If the food source is within about 50 m of the hive, a simple dance called the round dance—rapid movement in little circles, in both clockwise and counterclockwise directions—is sufficient. Outgoing foragers then know that the source is close and they search at random in the vicinity of the hive until they find the source.
If the food source is farther away, the incoming forager does a more complex dance, the waggle dance. The steps of the dance are illustrated in Fig. 45.15. The forager moves quickly back and forth (“waggles”) as it moves forward, circles back and repeats the waggle, then circles back the other way, and so on. This dance usually goes on for several minutes and may take much longer. The distance to the food source is conveyed by the length of time it takes to waggle up the middle of the circle. The direction, with reference to the position of the sun, is conveyed by the angle of the line of the waggle run: If the food source is in a direct line with the sun, the dancer’s waggle line points straight up; if food is at an angle of 45 degrees to the left of the sun, the dancer’s line is 45 degrees to the left of vertical. Short of human language, the bees’ dance language is among the natural world’s most extraordinary and effective methods of communication.