45.6 SOCIAL BEHAVIOR

Returning to Tinbergen’s questions, we can consider further the adaptive function and evolutionary history of animal behavior. The evolutionary advantages of most behaviors are readily apparent. The imprinting of Lorenz’s goslings on the first moving object they encountered makes sense when you consider that that object is usually their parent (not Konrad Lorenz!) and that sticking close to their parent is an excellent survival strategy in a world filled with predators.
Consider cooperative hunting seen in killer whales. Killer whales participate in a behavior called wave washing, in which a pod of whales collectively creates waves to wash a prey species—typically a seal—off an iceberg (Fig. 45.16). This behavior is collaborative, and it is clear why each individual participates. By itself, a single killer whale is incapable of generating a big enough wave, but, through cooperation, an individual can contribute to prey capture and gain some portion of the prey.
Sometimes, however, how a behavior enhances survival and reproduction is less clear. Perhaps the most prominent examples of this kind of behavior are sacrifices made by individuals apparently for the good of others. Formally, we describe an act of self-sacrifice as altruistic. Such acts seem to fly in the face of the natural selection, which posits that every individual competes for resources and mates to maximize its genetic contribution to the next generation. This contradiction is particularly stark in the case of species like honeybees: Honeybee workers, which are females, do not reproduce but instead assist the reproduction of the queen. Darwin confessed that such behaviors caused him sleepless nights. How, Darwin wondered, could altruism arise?
45.6.1 Group selection is a weak explanation of altruistic behavior.
Darwin himself suggested a solution to the altruism problem, a solution that today we call group selection. The idea is that natural selection operating on individuals is a less powerful force than another form of selection that operates on groups. Consider two groups, one with individuals that are altruistic, helping one another out, and another with individuals that act entirely selfishly. Darwin argued that, in a conflict between the two groups, the first group would beat the second because the first group works better as a unit. Here, then, we see that selection in favor of altruism that benefits the group has trumped selection in favor of selfish behavior.
Although an attractive idea, group selection is probably not important in the evolution of altruism. Altruism under group selection is typically not an evolutionarily stable strategy, meaning that this kind of behavior can be readily driven to extinction by an alternative strategy. It used to be thought that lemmings, a species of Scandinavian rodent, would collectively kill themselves by plunging into the ocean when the population became too large to sustain. It was claimed that the lemmings were altruistic: Their self-sacrificial act would ensure that the population did not exhaust available resources, preventing a population crash.
This is a nice story, but untrue. Lemmings do not kill themselves for the good of the species. An individual that “cheats” by not performing the altruistic act will survive and breed, and its altruistic companions will not. Now consider that this behavior—to kill oneself or not—is genetically encoded. A mutation that arises in a population that causes the individual to “cheat” and not kill itself will spread rapidly through the population by natural selection until, after many generations, it has completely replaced the altruistic allele that causes the individual to kill itself. The lemming behavior, then, is not an evolutionarily stable strategy because natural selection favors an alternative strategy.
This description is oversimplified because there is unlikely to be a simple genetic basis to traits such as these, but the point stands: Group selection is in general not an evolutionarily stable strategy because it can readily be overthrown by “selfish” strategies.
Question Quick Check 4
lDDGwjPw8LAlgih8oLKAmj5sYtjOhy7XKprPfJ+UqI7dhnWwexf5cK4lB+gtiBVsxbkGXoAT0eQrjA53U/4nI0I5iamwFlcsGnM/lpEAFAjFhZhJQ1nzDAzjR09j1VPvCaMZORkdJohBr6oYuNZ0p+yTFDZ93fuZ9LyXpYA8nKo+x+GpS07TPx3n57dUURO7spf1yg==45.6.2 Reciprocal altruism is one way that altruism can evolve.
If group selection does not provide a strong explanation for the evolution of altruism, how can we explain this behavior? Reciprocal altruism, whereby individuals exchange favors, is one way that altruism can evolve. This idea is perhaps best summarized as “You scratch my back, and I’ll scratch yours.” For this kind of behavior to work, individuals have to be able to recognize each other and remember previous interactions. If I did you a favor, I need to be able to recall who you are so that I can be sure that I get the return I expect.
A famous example of reciprocal altruism comes from an unlikely source—vampire bats. Adult female vampire bats live in groups of 8 to 12. Individuals have been recorded as living as long as 18 years, and in one study, two tagged females shared the same roost for 12 years. As is the case for most mammals, young males disperse from their natal group, but females remain. Thus, the female groups are composed partly of kin, although not entirely, since they are occasionally joined by unrelated females. In addition, because they have long-term associations, there is opportunity for reciprocal interactions.
Feeding for these bats is a “boom or bust” activity—either a bat has found and successfully fed on a blood source (boom), or a bat has failed to feed (bust). As a result, bats are often either overfed or underfed. Underfed bats, in turn, are at risk of dying by starvation. Bats returning from a successful feeding expedition often regurgitate blood to unsuccessful ones, on the expectation that the favor will be returned at a future date.
45.6.3 Kin selection is based on the idea that it is possible to contribute genetically to future generations by helping close relatives.
Only in the 1960s did we come to understand that the relatedness of participating individuals has played an important role in the evolution of altruism. This understanding lay in the insight that there are at least two ways to contribute genetically to the next generation: Either an individual can produce its own offspring, or it can produce no offspring and help close relatives reproduce instead. The latter strategy can become established in a population through a process called kin selection, a form of natural selection that favors the spread of alleles that promote behaviors that help close relatives, or kin.
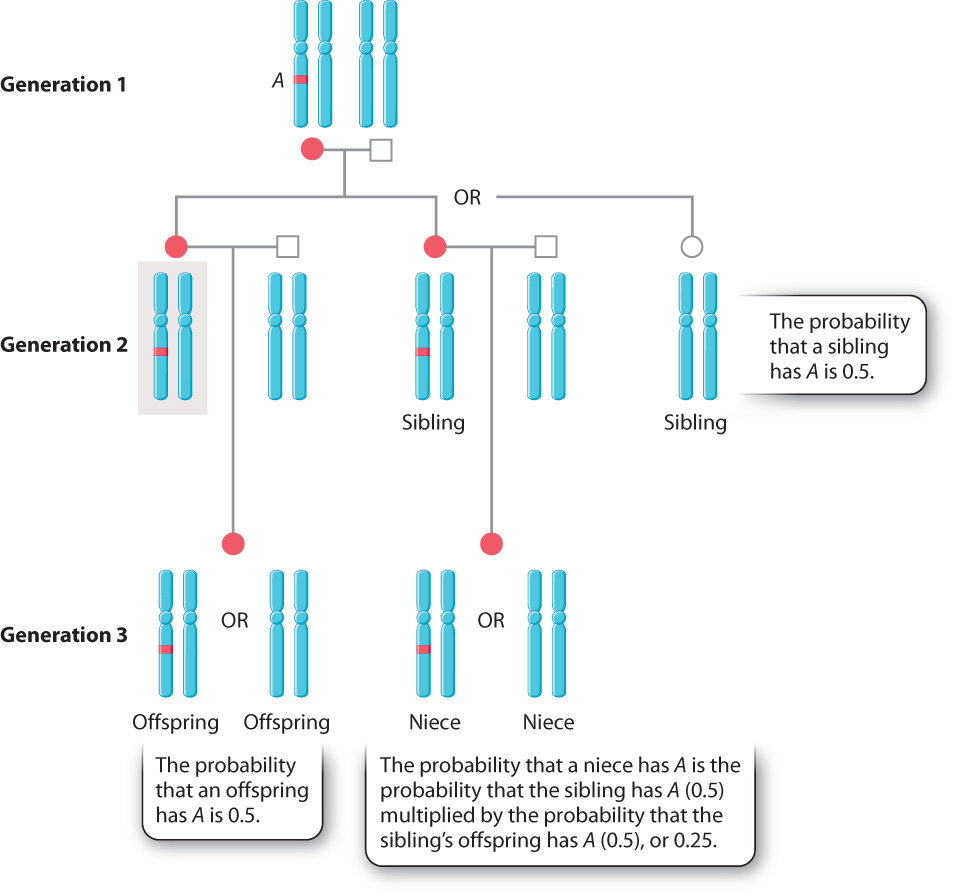
What relationships qualify as kin? Consider an animal (gray shading in the figure) that has an allele A at a particular locus (Fig. 45.17). If the animal reproduces, the probability of each offspring inheriting an A allele is 0.5. But what if the animal does not reproduce? According to Mendel’s laws, there is a 50% chance that the animal’s parents passed on the same A allele to the animal’s sibling (Chapter 16). If the sibling does, in fact, inherit allele A, the probability that it passes allele A to each of its offspring is also 0.5. Given the two probabilities (that the sibling has an A, and that the allele passed on to an offspring), the probability that the animal’s niece or nephew has A is 0.25. In other words, a diploid animal (such as humans) is twice as closely related to its own offspring than to its nephew or niece. Therefore, if the animal does not reproduce in order to help its sibling have two offspring, the animal will make the same net genetic contribution to the next generation as it would having one of its own offspring.
Question Quick Check 5
6VncWTNWFaHSIKmrfS5FvZwH8eoTTzuERH0+SHjbQnq10Ln3ANn+7hR0v4Xdu6ByVpFVCnnGpdml82nXtSXdSQEpOYm6DJHDi1z5yA==This logic was formalized in 1964 in a famous paper by British evolutionary biologist William D. Hamilton in terms of three key variables: B, the benefit of the behavior to the recipient (how many individuals does the behavior save?); C, the cost of the behavior to the donor (how many offspring are lost?); and r, the degree of relatedness between the recipient and donor. Hamilton asserted that as long as rB exceeds C, altruism can evolve.
How in practice does kin selection work? Let’s start with an extreme case of relatedness: clonal organisms that are genetically identical to each other (meaning that the probability of a sibling having allele A is 1). Many aphid species have a phase of their life cycle in which a “mother” aphid reproduces parthenogenetically to produce a small colony of clones of herself (Chapter 42). Any aphid in the colony is capable of reproducing in this way, but in a case of reproductive altruism, some individuals do not reproduce at all. They are “soldier” aphids, who defend the colony against predators, typically other insects. As specialized defenders, they do not have the ability to reproduce. In this case, the Hamilton calculation is simple. By helping their clone-mates survive, the sterile soldier aphids are ensuring that their own genes survive to be passed on to the next generation.
The most famous cases of reproductive altruism occur in the insect group Hymenoptera, which includes ants, bees, and wasps. Many species of Hymenoptera are eusocial, meaning that they have overlapping generations in a nest, cooperative care of the young, and clear and consistent division of labor between reproducers (the queen of a honeybee colony) and nonreproducers (the workers). Often called social insects, these species are one of evolution’s most extraordinary success stories.
What is it about Hymenoptera that predisposed them to eusociality? E. O. Wilson argues that natural selection has strongly favored group behavior in these insects. They typically receive group benefits from living in and defending a nest. For example, they can easily communicate with one another about the location of food resources such as nectar.
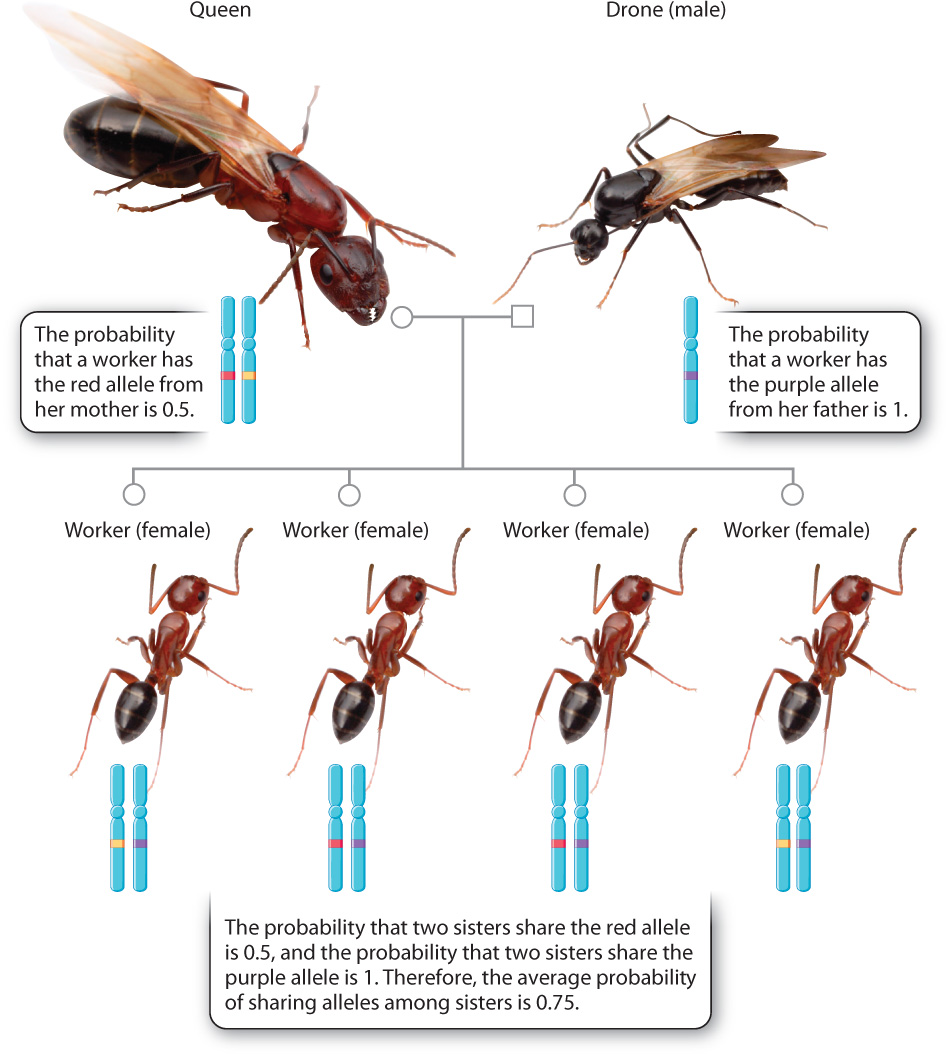
William Hamilton also noted that most Hymenoptera have an unusual mode of sex determination: Females (the queen and workers) are diploid, whereas males (drones) are haploid, produced from unfertilized eggs. As a result, the degree of relatedness, r, is higher for the sister–sister relationship (0.75) than for the mother–daughter one (0.5), as shown in Fig. 45.18. Put another way, a female is more closely related to her sister than she is to her daughter. Therefore, it makes more evolutionary sense, in terms of genetic representation in the next generation, for a worker to help her mother, the queen, to produce more sisters than for her to produce her own offspring.
This form of social organization can result in a huge colony of related individuals all working collectively, much as different cells work collectively within a body. Often, we see specialization into castes: Some ants are soldiers, defending the colony, others are foragers, and so on. If we extend the analogy of individuals acting like cells in a body, the multiple castes are equivalent to organs, all coordinating and working together. The term “superorganism” is sometimes used to describe a single social insect colony.