47.2 ANTAGONISTIC INTERACTIONS BETWEEN SPECIES

The realized niche of tree frogs compared with their fundamental niche makes it clear that interactions with other species play key roles in the distribution and abundance of populations. Close interactions between species that have evolved over long periods of time are called symbioses (Chapter 26). Many of these interactions enhance the reproduction and population growth of both ecological partners, like chocolate trees and midges (Fig. 47.3). These mutually beneficial interactions are called mutualisms. Many other interactions are one sided: One organism actually consumes another or competes with it for food or some other resource (Fig. 47.4). Such one-sided interactions, in which at least one participant loses more than it gains, are called antagonisms.
All interactions combine costs (often in physical resources) and benefits (usually increases in reproduction), and the ratio of costs to benefits for each of the participants reveals whether an overall interaction is mutualistic or antagonistic. Some interactions are direct, where species physically interact (such as plants and their pollinators). Others are indirect, where they influence one another through competition for a shared resource, such as food. Virtually every organism is involved in both mutualisms and antagonisms with other species—think of plants and their pollinators or predators and their prey. These interactions often narrow the geographic distributions of species within the larger boundaries of their physical requirements, and so help shape their realized niches.

47.2.1 Limited resources foster competition.
Whenever there are fewer resources than individuals seeking them, there can be competition. Competition is an interaction in which the use of a mutually needed resource by one individual or group of individuals lowers the availability of the resource for another individual or group. Different species of sea anemones compete for space in rocky shallows, swallows compete with one another for nesting holes in tree or cliffs, and many different species of dung beetles compete for animal droppings (some even cling to monkeys, waiting for feces to emerge). Male birds, mammals, insects, and other animals commonly compete for female mates.
In Chapter 46, we saw that in many populations the tension between intrinsic rate of reproduction and environmental carrying capacity results in intraspecific competition—competition within a species. The increase in competition between individuals of a species that accompanies increased population density is a main reason why population growth slows as the species’ environment approaches its carrying capacity. As noted earlier in this chapter, competition can also occur between individuals of different species, in which case it is interspecific competition. Whether competition occurs between individuals of one species or two different ones, it is a lose–lose situation: Both sides spend energy they would not in the absence of the other. Indeed, Darwin recognized that the “struggle for existence,” as he called it, is a primary driver for natural selection, leading to the evolution of traits that aid survival and reproduction by reducing competition in one way or another.
47.2.2 Competition promotes niche divergence.
In general, when two co-occurring species overlap strongly in resource use, one will either become extinct in that place or change its resource use. For example, coyotes and foxes eat the same small prey, mice and rabbits, but when they meet, a coyote will waste energy by chasing the fox and of course the fox loses energy by running away from the coyote. In regions where foxes and coyotes occur together, the foxes avoid interactions with the larger coyotes by leaving the area, placing their dens in locations where coyotes do not occur. This result of such an antagonistic interaction, in which one species is prevented from occupying a particular habitat or niche, is called competitive exclusion. This is another way that a fundamental niche can be reduced to a smaller realized niche. Over time, competitive exclusion leads to resource partitioning, as we saw with the Anolis lizards (Chapter 46).
Species can compete with each other directly or indirectly. For example, some trees compete with other trees directly by producing toxins in the soil around them that the seedlings of other species cannot tolerate, and corals may compete by stinging overlapping colonies. Some species compete with others indirectly by interfering with the resources used by a different species. For example, diatom algae make skeletons of silica (SiO2), depleting this substance in the surface waters of the world’s oceans. Radiolaria also make skeletons of silica, and for the past 65 million years, radiolarians have evolved silica-saving modifications of their skeletons in response to the increasing sequestration of SiO2 by diatoms.
47.2.3 Species compete for resources other than food.
Food is the resource we most often think of as the limiting factor for population growth and reproduction, but many species just require a place to stand, so to speak, or suitable conditions for a nest. Barnacle and mussel larvae compete for a place to attach on submerged rocks. Their food, tiny plankton, is almost always abundant and therefore is not as likely to limit population growth as attachment space is.
Similarly, plants compete for space in fields and forests so they can collect sunlight and obtain water and nutrients from the soil. Sunlight cannot itself be exhausted by use, but room to collect sunlight on the ground below can be filled. Nesting sites for birds that lay their eggs in tree hollows or on cliff faces can also be strongly limited by the availability of space, as can the settling of corals on reef surfaces (Fig. 47.5).

47.2.4 Predators and parasites can limit prey population size, minimizing competition.
Predation is another form of antagonism, as the predator benefits at the expense of the prey. Predators can limit the population sizes of their prey, preventing prey populations from increasing to the level where competitive exclusion occurs. We have seen that when species overlap in resource use and their populations increase to the point where they compete for resources, they must be separated in space or time or risk local extinction through competitive exclusion when resources become limited.
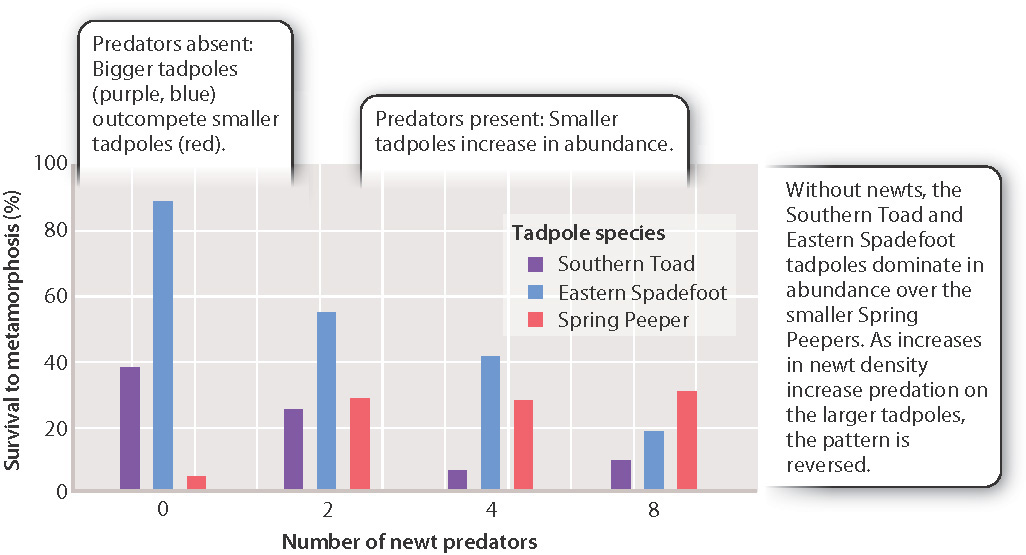
If, however, populations do not rise to densities at which resources are limiting, competition is reduced and species can overlap in niches without excluding one another. For example, in ponds throughout the southern United States, tadpoles of the large Southern Toad, the Eastern Spadefoot, and the tiny Spring Peeper all use the same food source. In the presence of the two larger species, Peeper tadpoles compete poorly for food (Fig. 47.6). As a result, they have low survival rates, and those individuals that do make it to maturity are often undersized because as tadpoles they did not have access to adequate nutrition. In the presence of a predatory Red-Spotted Newt, however, the tables are turned. The newt prefers to eat the larger tadpole species, and so prevents them from becoming sufficiently abundant to outcompete the smaller Peeper tadpoles. The Peeper tadpoles, then, have the highest survival rates. With competition for food reduced, these small tadpoles grow to their maximal body size, and become the most abundant tadpole species in the ponds.
Parasites can also limit the population size of their host, keeping numbers well below the carrying capacity of the environment. An extreme example is the fungus that infects the American chestnut, a tree once dominant in eastern North American forests. The fungus attacks the vascular system of young chestnut trees, which usually succumb when they reach 2 to 3 m in height. Today, there are only a few isolated patches of large chestnut trees in Vermont and a handful of other places that have escaped the fungus. As chestnut populations have declined, populations of oak, beech, and other trees that grow in the same forest have increased.