Sex chromosomes
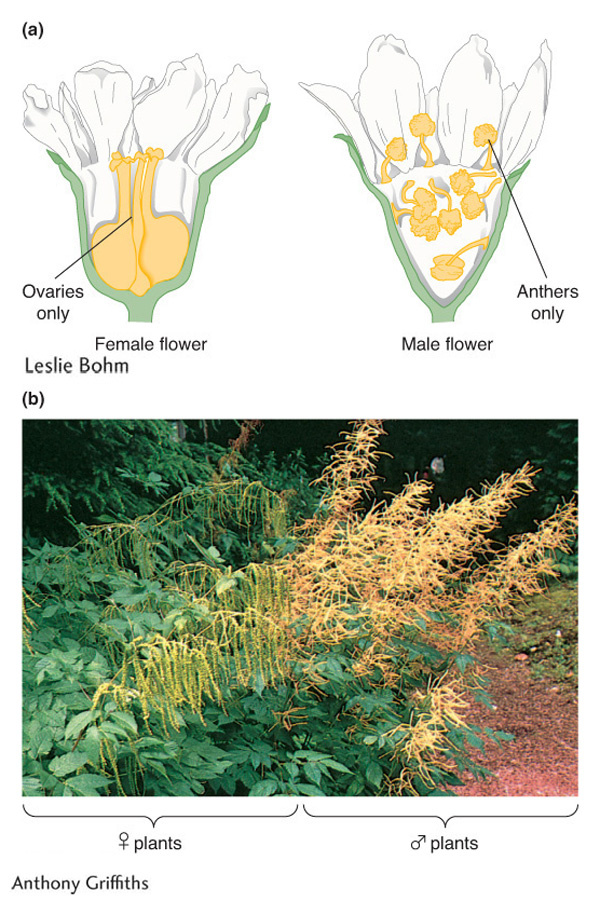
Figure 2-14: Male and female plants
Figure 2-14: Examples of two dioecious plant species are (a) Osmaronia dioica and (b) Aruncus dioicus.
[(a) Leslie Bohm; (b) Anthony Griffiths]
Most animals and many plants show sexual dimorphism; in other words, individuals are either male or female. In most of these cases, sex is determined by a special pair of sex chromosomes. Let’s look at humans as an example. Human body cells have 46 chromosomes: 22 homologous pairs of autosomes plus 2 sex chromosomes. Females have a pair of identical sex chromosomes called the X chromosomes. Males have a nonidentical pair, consisting of one X and one Y. The Y chromosome is considerably shorter than the X. Hence, if we let A represent autosomal chromosomes, we can write
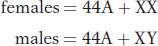
At meiosis in females, the two X chromosomes pair and segregate like autosomes, and so each egg receives one X chromosome. Hence, with regard to sex chromosomes, the gametes are of only one type and the female is said to be the homogametic sex. At meiosis in males, the X and the Y chromosomes pair over a short region, which ensures that the X and Y separate so that there are two types of sperm, half with an X and the other half with a Y. Therefore, the male is called the heterogametic sex.
The inheritance patterns of genes on the sex chromosomes are different from those of autosomal genes. Sex-chromosome inheritance patterns were first investigated in the early 1900s in the laboratory of the great geneticist Thomas Hunt Morgan, using the fruit fly Drosophila melanogaster (see the Model Organism box). This insect has been one of the most important research organisms in genetics; its short, simple life cycle contributes to its usefulness in this regard. Fruit flies have three pairs of autosomes plus a pair of sex chromosomes, again referred to as X and Y. As in mammals, Drosophila females have the constitution XX and males are XY. However, the mechanism of sex determination in Drosophila differs from that in mammals. In Drosophila, the number of X chromosomes in relation to the autosomes determines sex: two X’s result in a female, and one X results in a male. In mammals, the presence of the Y chromosome determines maleness and the absence of a Y determines femaleness. However, it is important to note that, despite this somewhat different basis for sex determination, the single-gene inheritance patterns of genes on the sex chromosomes are remarkably similar in Drosophila and mammals.
Vascular plants show a variety of sexual arrangements. Dioecious species are those showing animal-like sexual dimorphism, with female plants bearing flowers containing only ovaries and male plants bearing flowers containing only anthers (Figure 2-14). Some, but not all, dioecious plants have a nonidentical pair of chromosomes associated with (and almost certainly determining) the sex of the plant. Of the species with nonidentical sex chromosomes, a large proportion have an XY system. For example, the dioecious plant Melandrium album has 22 chromosomes per cell: 20 autosomes plus 2 sex chromosomes, with XX females and XY males. Other dioecious plants have no visibly different pair of chromosomes; they may still have sex chromosomes but not visibly distinguishable types.
Sex-linked patterns of inheritance
Cytogeneticists divide the X and Y chromosomes into homologous and differential regions. Again, let’s use humans as an example (Figure 2-15). The differential regions, which contain most of the genes, have no counterparts on the other sex chromosome. Hence, in males, the genes in the differential regions are said to be hemizygous (“half zygous”). The differential region of the X chromosome contains many hundreds of genes; most of these genes do not take part in sexual function, and they influence a great range of human properties. The Y chromosome contains only a few dozen genes. Some of these genes have counterparts on the X chromosome, but most do not. The latter type take part in male sexual function. One of these genes, SRY, determines maleness itself. Several other genes are specific for sperm production in males.
Figure 2-15: Human sex chromosomes
Figure 2-15: Human sex chromosomes contain a differential region and two pairing regions. The regions were located by observing where the chromosomes paired up in meiosis and where they did not.
In general, genes in the differential regions are said to show inheritance patterns called sex linkage. Mutant alleles in the differential region of the X chromosome show a single-gene inheritance pattern called X linkage. Mutant alleles of the few genes in the differential region of the Y chromosome show Y linkage. A gene that is sex linked can show phenotypic ratios that are different in each sex. In this respect, sex-linked inheritance patterns contrast with the inheritance patterns of genes in the autosomes, which are the same in each sex. If the genomic location of a gene is unknown, a sex-linked inheritance pattern indicates that the gene lies on a sex chromosome.
The human X and Y chromosomes have two short homologous regions, one at each end (see Figure 2-15). In the sense that these regions are homologous, they are autosomal-like, and so they are called pseudoautosomal regions 1 and 2. One or both of these regions pairs in meiosis and undergoes crossing over (see Chapter 4 for details of crossing over). For this reason, the X and the Y chromosomes can act as a pair and segregate into equal numbers of sperm.
X-linked inheritance
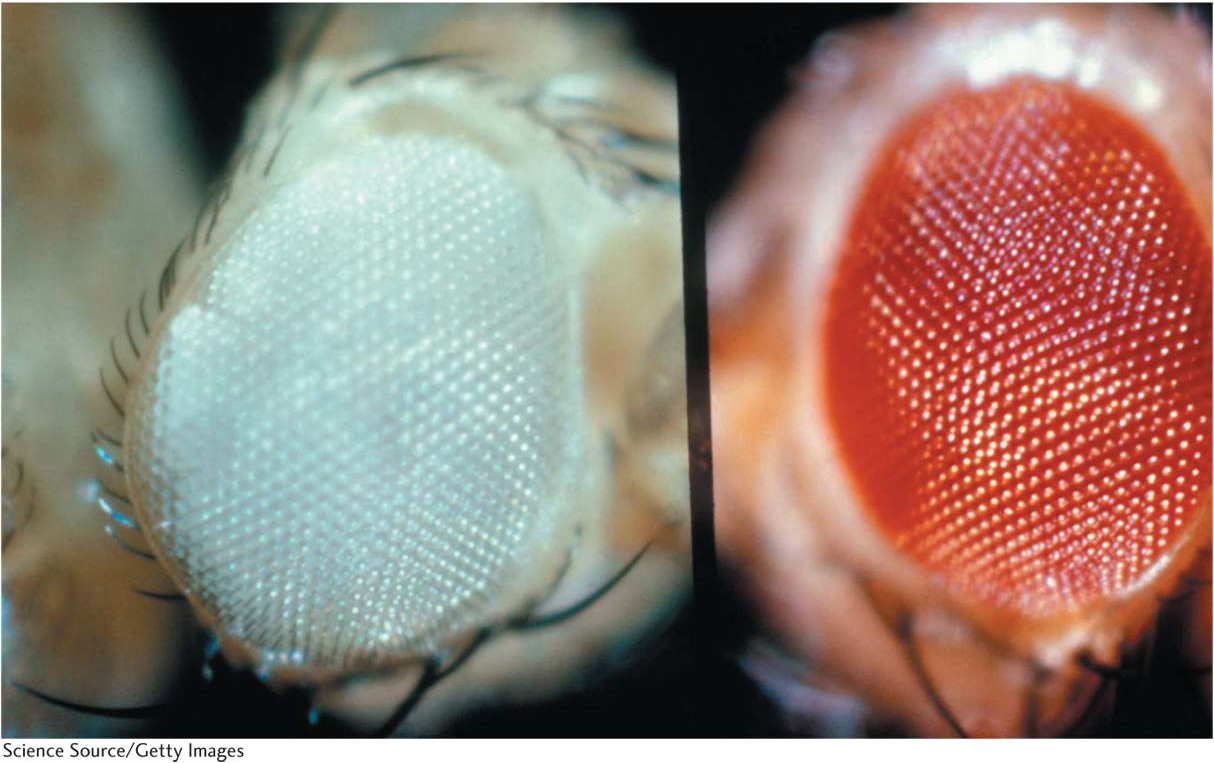
Figure 2-16: White-eyed and red-eyed Drosophila
Figure 2-16: The red-eyed fly is wild type, and the white-eyed fly is a mutant.
[Science Source/Getty Images.]
For our first example of X linkage, we turn to eye color in Drosophila. The wild-type eye color of Drosophila is dull red, but pure lines with white eyes are available (Figure 2-16). This phenotypic difference is determined by two alleles of a gene located on the differential region of the X chromosome. The mutant allele in the present case is w for white eyes (the lowercase letter indicates that the allele is recessive), and the corresponding wild-type allele is w+. When white-eyed males are crossed with red-eyed females, all the F1 progeny have red eyes, suggesting that the allele for white eyes is recessive. Crossing these red-eyed F1 males and females produces a 3:1 F2 ratio of red-eyed to white-eyed flies, but all the white-eyed flies are males. This inheritance pattern, which shows a clear difference between the sexes, is explained in Figure 2-17. The basis of the inheritance pattern is that all the F1 flies receive a wild-type allele from their mothers, but the F1 females also receive a white-eye allele from their fathers. Hence, all F1 females are heterozygous wild type (w+/w), and the F1 males are hemizygous wild type (w+). The F1 females pass on the white-eye allele to half their sons, who express it, and to half their daughters, who do not express it, because they must inherit the wild-type allele from their fathers.
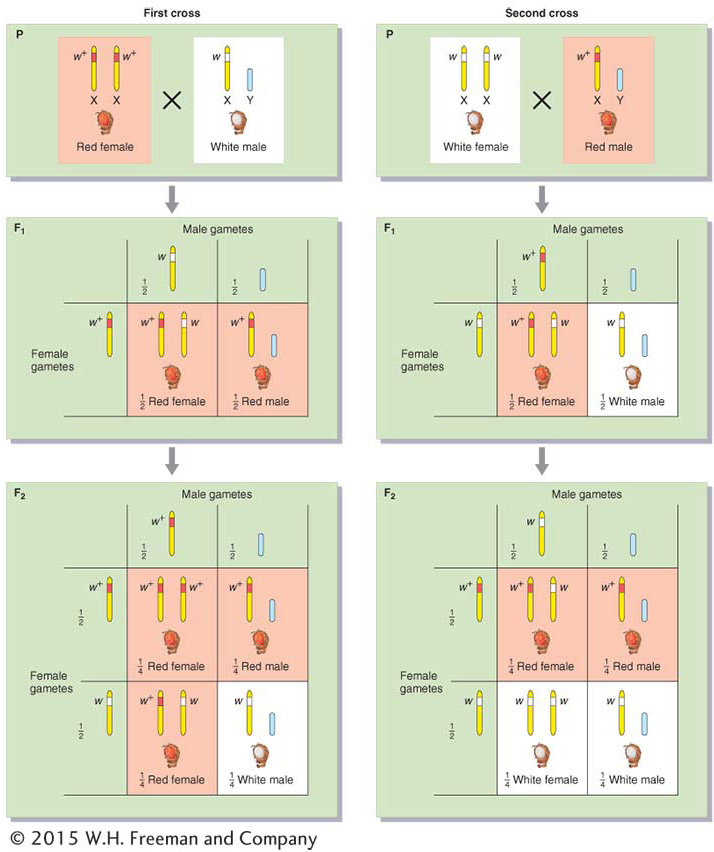
Figure 2-17: An example of X-linked inheritance
Figure 2-17: Reciprocal crosses between red-eyed (red) and white-eyed (white) Drosophila give different results. The alleles are X linked, and the inheritance of the X chromosome explains the phenotypic ratios observed, which are different from those of autosomal genes. (In Drosophila and many other experimental systems, a superscript plus sign is used to designate the normal, or wild-type, allele. Here, w+ encodes red eyes and w encodes white eyes.)
Drosophila
Drosophila melanogaster was one of the first model organisms to be used in genetics. It is readily available from ripe fruit, has a short life cycle, and is simple to culture and cross. Sex is determined by X and Y sex chromosomes (XX = female, XY = male), and males and females are easily distinguished. Mutant phenotypes regularly arise in lab populations, and their frequency can be increased by treatment with mutagenic radiation or chemicals. It is a diploid organism, with four pairs of homologous chromosomes (2n = 8). In salivary glands and certain other tissues, multiple rounds of DNA replication without chromosomal division result in “giant chromosomes,” each with a unique banding pattern that provides geneticists with landmarks for the study of chromosome mapping and rearrangement. There are many species and races of Drosophila, which have been important raw material for the study of evolution.
Time flies like an arrow; fruit flies like a banana. (Groucho Marx)

Drosophila melanogaster, the common fruit flt.
[© blickwinkel/Alamy.]
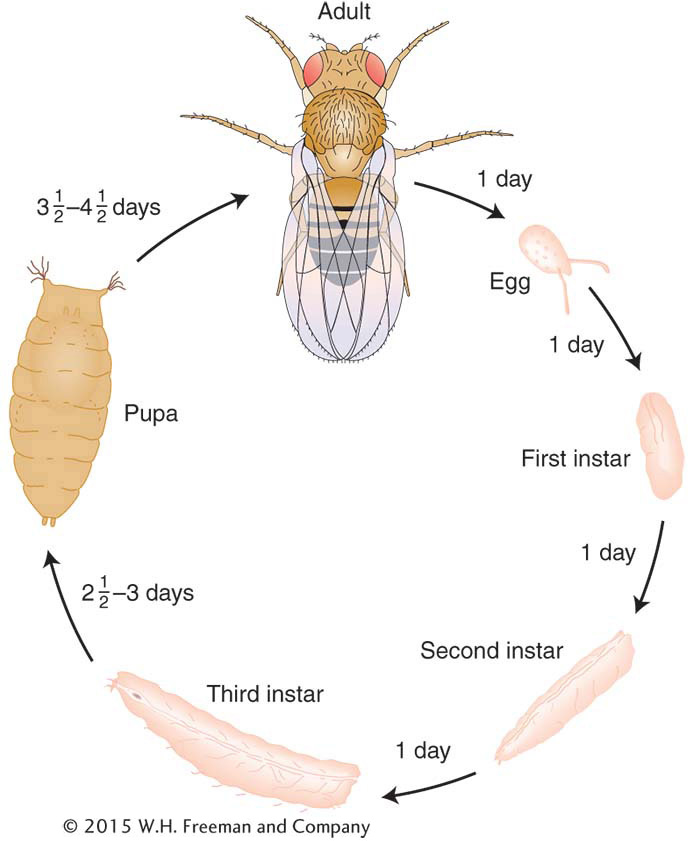
Life cycle of Drosophila melanogaster.
The reciprocal cross gives a different result; that is, the cross between white-eyed females and red-eyed males gives an F1 in which all the females are red eyed but all the males are white eyed. In this case, every female inherited the dominant w+ allele from the father’s X chromosome, whereas every male inherited the recessive w allele from its mother. The F2 consists of one-half red-eyed and one-half white-eyed flies of both sexes. Hence, in sex linkage, we see examples not only of different ratios in different sexes, but also of differences between reciprocal crosses.
Note that Drosophila eye color has nothing to do with sex determination, and so we have an illustration of the principle that genes on the sex chromosomes are not necessarily related to sexual function. The same is true in humans: in the discussion of pedigree analysis later in this chapter, we shall see many X-linked genes, yet few could be construed as being connected to sexual function.
The abnormal allele associated with white eye color in Drosophila is recessive, but abnormal alleles of genes on the X chromosome that are dominant also arise, such as the Drosophila mutant hairy wing (Hw). In such cases, the wild-type allele (Hw+) is recessive. The dominant abnormal alleles show the inheritance pattern corresponding to that of the wild-type allele for red eyes in the preceding example. The ratios obtained are the same.
KEY CONCEPT
Sex-linked inheritance regularly shows different phenotypic ratios in the two sexes of progeny, as well as different ratios in reciprocal crosses.Historically, in the early decades of the twentieth century, the demonstration by Morgan of X-linked inheritance of white eyes in Drosophila was a key piece of evidence that suggested that genes are indeed located on chromosomes, because an inheritance pattern was correlated with one specific chromosome pair. The idea became known as “the chromosome theory of inheritance.” At that period in history, it had recently been shown that, in many organisms, sex is determined by an X and a Y chromosome and that, in males, these chromosomes segregate equally at meiosis to regenerate equal numbers of males and females in the next generation. Morgan recognized that the inheritance of alleles of the eye-color gene is exactly parallel to the inheritance of X chromosomes at meiosis; hence, the gene was likely to be on the X chromosome. The inheritance of white eyes was extended to Drosophila lines that had abnormal numbers of sex chromosomes. With the use of this novel situation, it was still possible to predict gene-inheritance patterns from the segregation of the abnormal chromosomes. That these predictions proved correct was a convincing test of the chromosome theory.
Other genetic analyses revealed that, in chickens and moths, sex-linked inheritance could be explained only if the female was the heterogametic sex. In these organisms, the female sex chromosomes were designated ZW and males were designated ZZ.