Arabidopsis thaliana
| Genetic "Vital Statistics" | |
|---|---|
|
Genome size: |
125 Mb |
|
Chromosomes: |
diploid, 5 autosomes (2n = 10) |
|
Number of genes: |
25,000 |
|
Percentage with human homologs: |
18% |
|
Average gene size: |
2 kb, 4 introns/gene |
|
Transposons: |
10% of the genome |
|
Genome sequenced in: |
2000 |
Arabidopsis thaliana

Key organism for studying:
Development
Gene expression and regulation
Plant genomics
Arabidopsis thaliana, a member of the Brassicaceae (cabbage) family of plants, is a relatively late arrival as a genetic model organism. Most work has been done in the past 20 years. It has no economic significance: it grows prolifically as a weed in many temperate parts of the world. However, because of its small size, short life cycle, and small genome, it has overtaken the more traditional genetic plant models such as corn and wheat and has become the dominant model for plant molecular genetics.

Special features
In comparison with other plants, Arabidopsis is small in regard to both its physical size and its genome size—
Genetic analysis
The analysis of Arabidopsis mutations through crossing relies on tried and true methods—
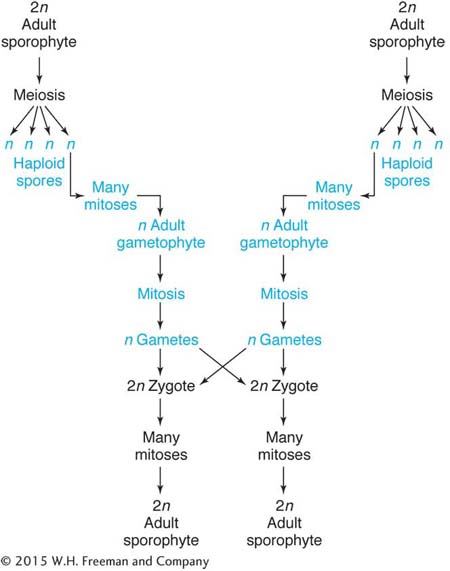
Life Cycle
Arabidopsis has the familiar plant life cycle, with a dominant diploid stage. A plant bears several flowers, each of which produces many seeds. Like many annual weeds, its life cycle is rapid: it takes only about 6 weeks for a planted seed to produce a new crop of seeds.
Total length of life cycle: 6 weeks
801

Techniques of Genetic Modification
|
Standard mutagenesis: |
|
|
Chemicals and radiation |
Random germ- |
|
T- |
Random tagged insertions |
|
Transgenesis: |
|
|
T- |
Random insertion |
|
Targeted gene knockouts: |
|
|
T- |
Random insertion; mutagenesis knockouts selected with PCR |
|
RNAi |
Mimics targeted knockout |
Genetic engineering
Transgenesis. Agrobacterium T-
Large collections of T-
Main contributions
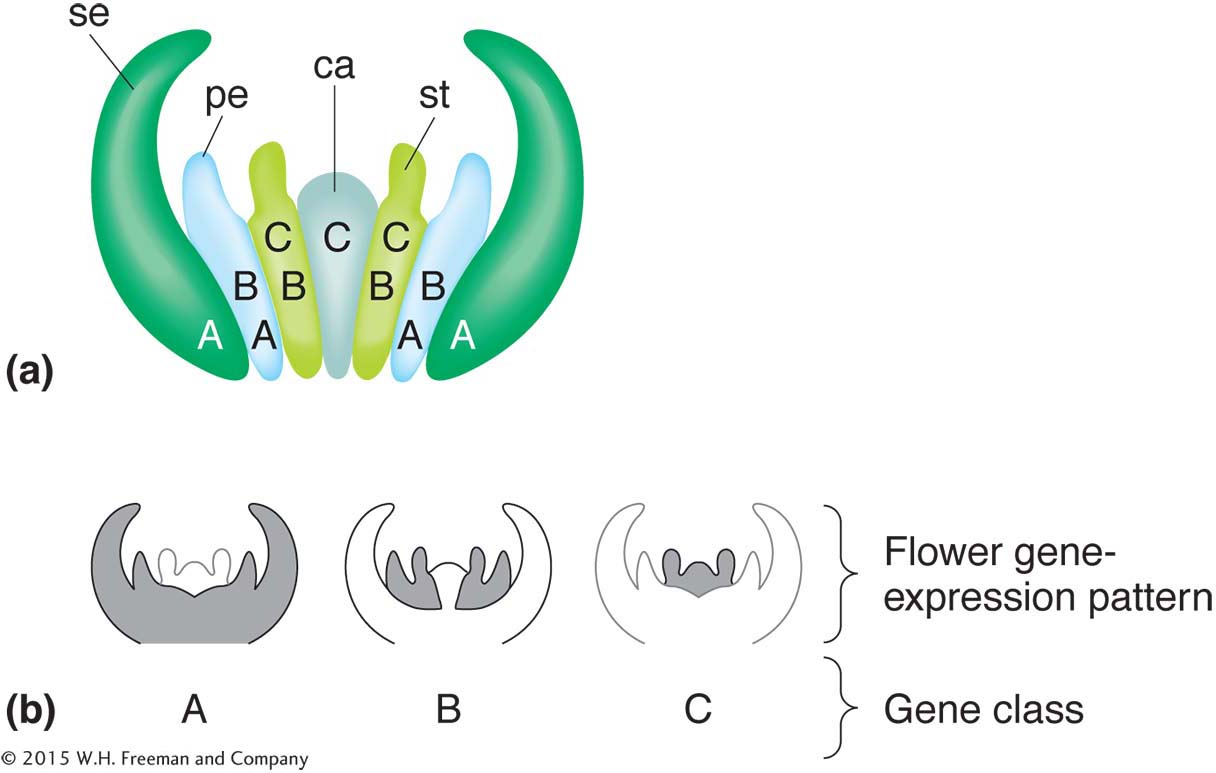
As the first plant genome to be sequenced, Arabidopsis has provided an important model for plant genome architecture and evolution. In addition, studies of Arabidopsis have made key contributions to our understanding of the genetic control of plant development. Geneticists have isolated homeotic mutations affecting flower development, for example. In such mutants, one type of floral part is replaced by another. Integration of the action of these mutants has led to an elegant model of flower-
Other areas of contribution
Environmental-
stress response Hormone control systems
802