15.1 Discovery of Transposable Elements in Maize
McClintock’s experiments: the Ds element
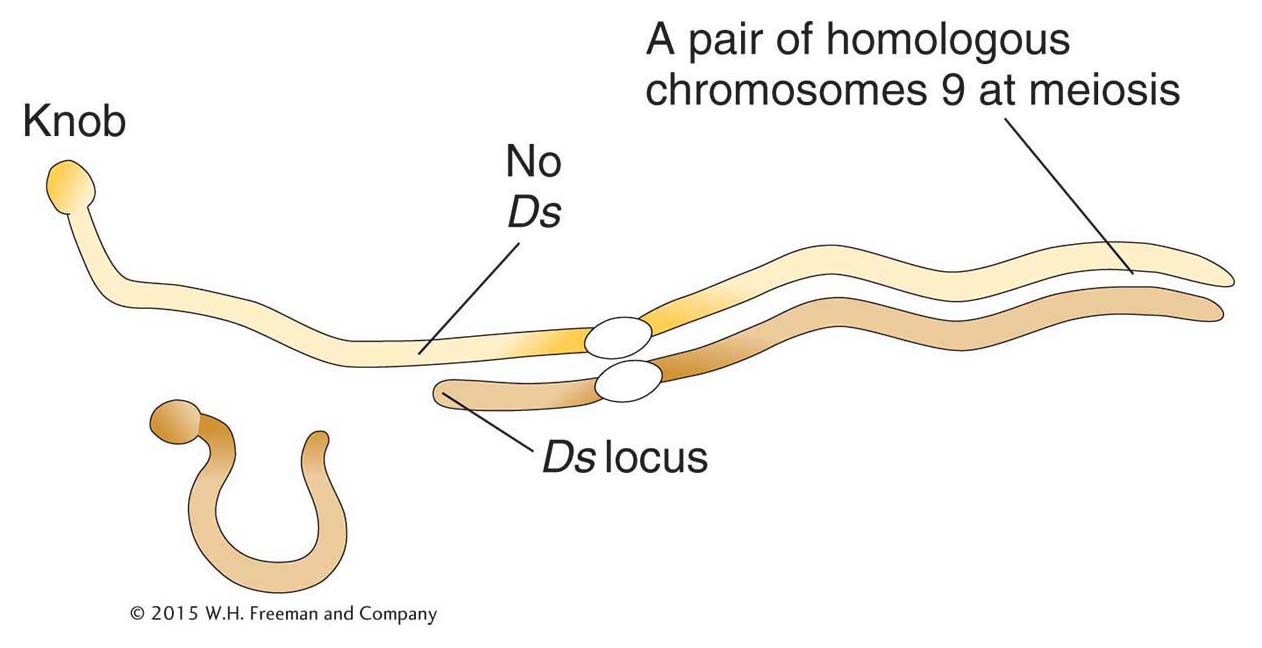
In the 1940s, Barbara McClintock made an astonishing discovery while studying the colored kernels of so-
McClintock began to suspect that Ac and Ds were actually mobile genetic elements when she found it impossible to map Ac. In some plants, it mapped to one position; in other plants of the same line, it mapped to different positions. As if this variable mapping were not enough of a curiosity, rare kernels with dramatically different phenotypes could be derived from the original strain that had frequent breaks in chromosome 9. One such phenotype was a rare colorless kernel containing pigmented spots.
Figure 15-3 compares the phenotype of the chromosome-
To explain the new derivatives, McClintock hypothesized that Ds had moved from a site near the centromere into the C gene located close to the telomeric end. In its new location, Ds prevents the expression of C. The inactivation of the C gene explains the colorless parts of the kernel, but what explains the appearance of the pigmented spots? The spotted kernel is an example of an unstable phenotype. McClintock concluded that such unstable phenotypes resulted from the movement or transposition of Ds away from the C gene. That is, the kernel begins development with a C gene that has been mutated by the insertion of Ds. However, in some cells of the kernel, Ds leaves the C gene, allowing the mutant phenotype to revert to wild type and produce pigment in the original cell and in all its mitotic descendants. There are big spots of color when Ds leaves the C gene early in kernel development (because there are more mitotic descendants), whereas there are small spots when Ds leaves the C gene later in kernel development. Unstable mutant phenotypes that revert to wild type are a clue to the participation of mobile elements.
550
Autonomous and nonautonomous elements
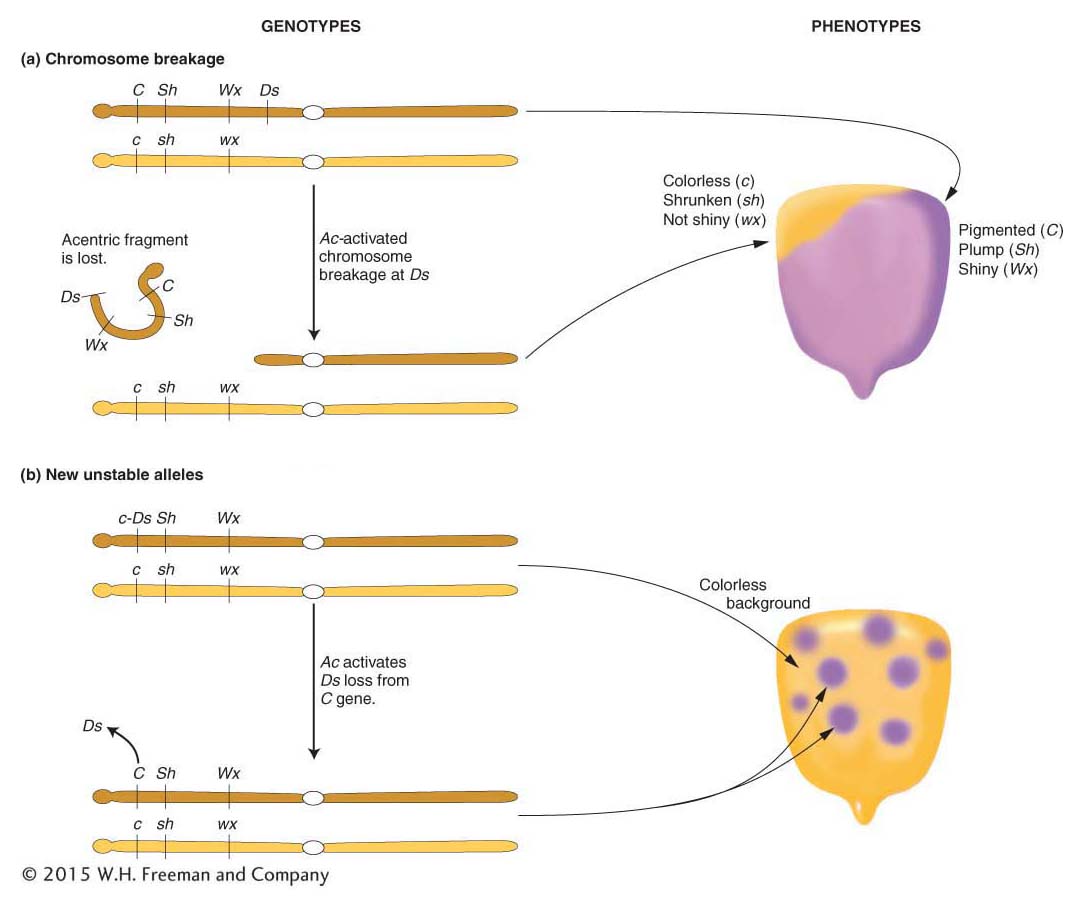
What is the relation between Ac and Ds? How do they interact with genes and chromosomes to produce these interesting and unusual phenotypes? These questions were answered by further genetic analysis. Interactions between Ds, Ac, and the pigment gene C are used as an example in Figure 15-4. There, Ds is shown as a piece of DNA that has inactivated the C gene by inserting into its coding region. The allele carrying the insert is called c-
Other strains were isolated in which the Ac element itself had inserted into the C gene [called c-
Several systems like Ac/Ds were found by McClintock and other geneticists working with maize. Two other systems are Dotted [(Dt), discovered by Marcus Rhoades] and Suppressor/mutator [(Spm), independently discovered by McClintock and Peter Peterson, who called it Enhancer/Inhibitor (En/In)]. In addition, as you will see in the sections that follow, elements with similar genetic behavior have been isolated from bacteria, plants, and animals.
551
Maize
Maize, also known as corn, is actually Zea mays, a member of the grass family. Grasses—
In the 1920s, Rollins A. Emerson set up a laboratory at Cornell University to study the genetics of corn traits, including kernel color, which were ideal for genetic analysis. In addition, the physical separation of male and female flowers into the tassel and ear, respectively, made controlled genetic crosses easy to accomplish. Among the outstanding geneticists attracted to the Emerson laboratory were Marcus Rhoades, Barbara McClintock, and George Beadle (see Chapter 6). Before the advent of molecular biology and the rise of microorganisms as model organisms, geneticists performed microscopic analyses of chromosomes and related their behavior to the segregation of traits. The large root-

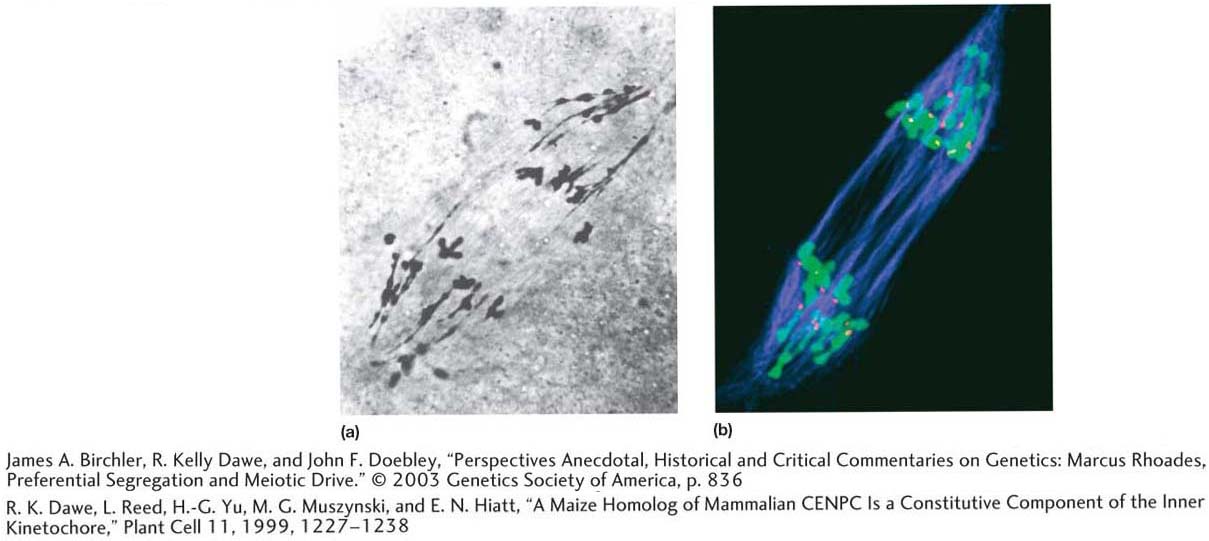
Maize still serves as a model genetic organism. Molecular biologists continue to exploit its beautiful pachytene chromosomes with new antibody probes (see photograph b below) and have used its wealth of genetically well-
552
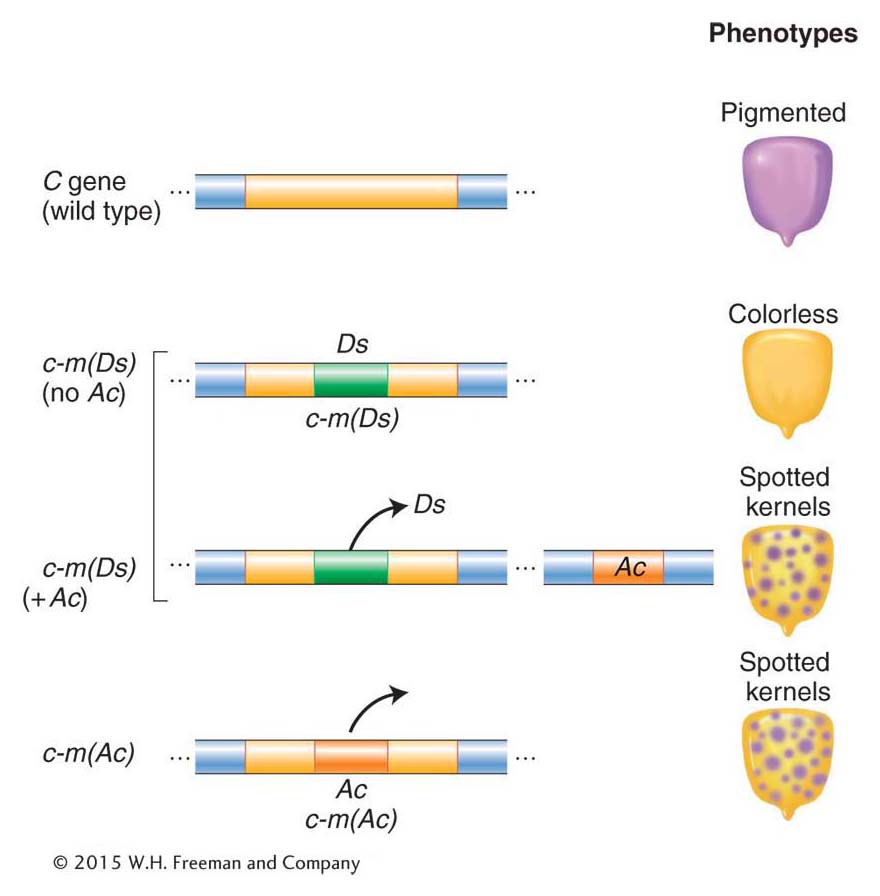
The common genetic behavior of these elements led geneticists to propose new categories for all the elements. Ac and elements with similar genetic properties are now called autonomous elements because they require no other elements for their mobility. Similarly, Ds and elements with similar genetic properties are called nonautonomous elements. An element family is composed of one or more autonomous elements and the nonautonomous members that can be mobilized. Autonomous elements encode the information necessary for their own movement and for the movement of unlinked nonautonomous elements in the genome. Because nonautonomous elements do not encode the functions necessary for their own movement, they cannot move unless an autonomous element from their family is present somewhere else in the genome.
Figure 15-5 shows an example of the effects of transposons in a rose.

KEY CONCEPT
Transposable elements in maize can inactivate a gene in which they reside, cause chromosome breaks, and transpose to new locations within the genome. Autonomous elements can perform these functions unaided; nonautonomous elements can transpose only with the help of an autonomous element elsewhere in the genome.Transposable elements: only in maize?
Although geneticists accepted McClintock’s discovery of transposable elements in maize, many were reluctant to consider the possibility that similar elements resided in the genomes of other organisms. Their existence in all organisms would imply that genomes are inherently unstable and dynamic. This view was inconsistent with the fact that the genetic maps of members of the same species were the same. After all, if genes can be genetically mapped to a precise chromosomal location, doesn’t this mapping indicate that they are not moving around?
553
Because McClintock was a highly respected geneticist, her results were not questioned. Rather, their relevance to other organisms was questioned by others who argued that maize is not a natural organism: it is a crop plant that is the product of human selection and domestication. This view was held by some until the 1960s, when the first transposable elements were isolated from the E. coli genome and studied at the DNA-