15-5 Variations in Cognitive Organization
No two brains are identical. Some differences are genetically determined; others result from plastic changes caused, for example, by experience and learning or epigenetic factors. Some brain differences are idiosyncratic (unique to a particular person); many others are systematic and common to whole categories of people. In this section, we consider two systematic variations in brain organization, those related to sex and handedness, and one idiosyncratic variation, the fascinating sensory phenomenon synesthesia.
Sex Differences in Cognitive Organization
The idea that men and women think differently probably originated with the first men and women. Science backs it up. Books, including one by Doreen Kimura (1999), present persuasive evidence for marked differences between men’s and women’s performance on many cognitive tests. As illustrated in Figure 15-15, paper-
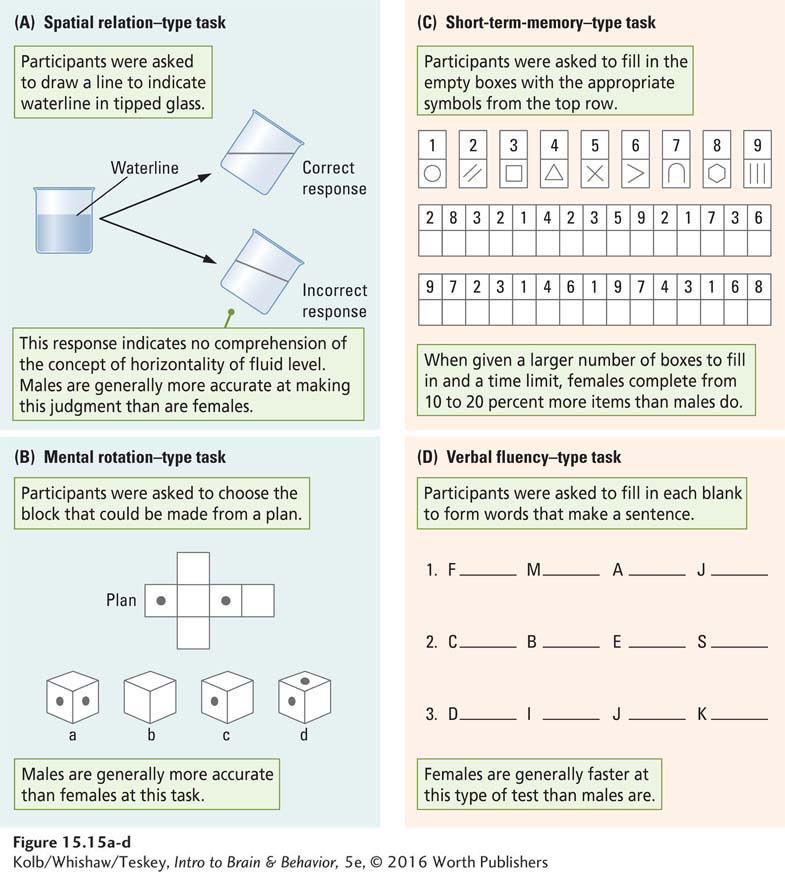
547
Neural Bases of Sex Differences
Considerable evidence points to sex differences, both in the brain’s gross cerebral structure and at a neuronal level. Jill Goldstein and her colleagues (2001) conducted a large MRI study of sexual dimorphism in the human brain. They found that women have larger volumes of dorsal prefrontal and associated paralimbic regions, whereas men have larger volumes of more ventral prefrontal regions (Figure 15-16). (Brain size is related to body size, and on average, male brains are bigger than female brains, so the investigators corrected for size.)
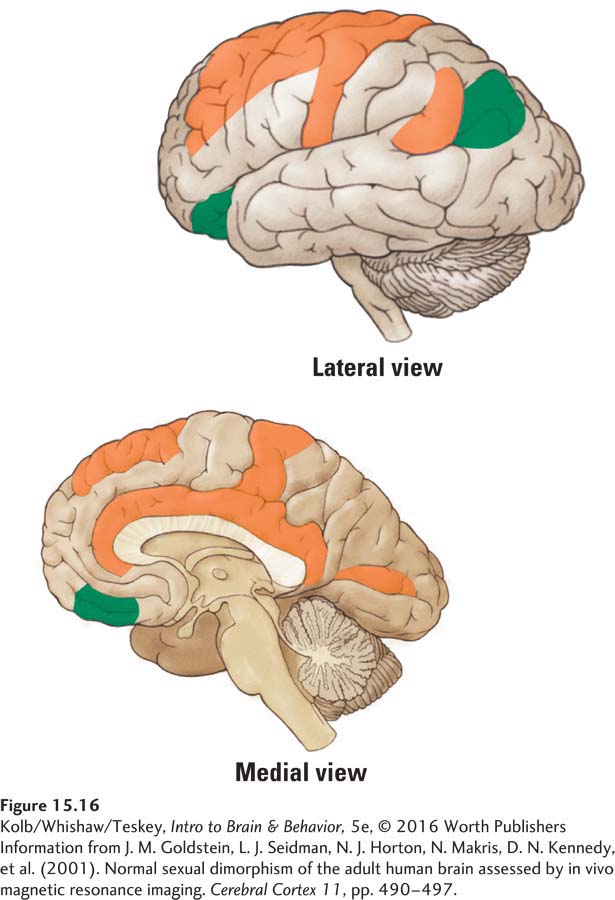
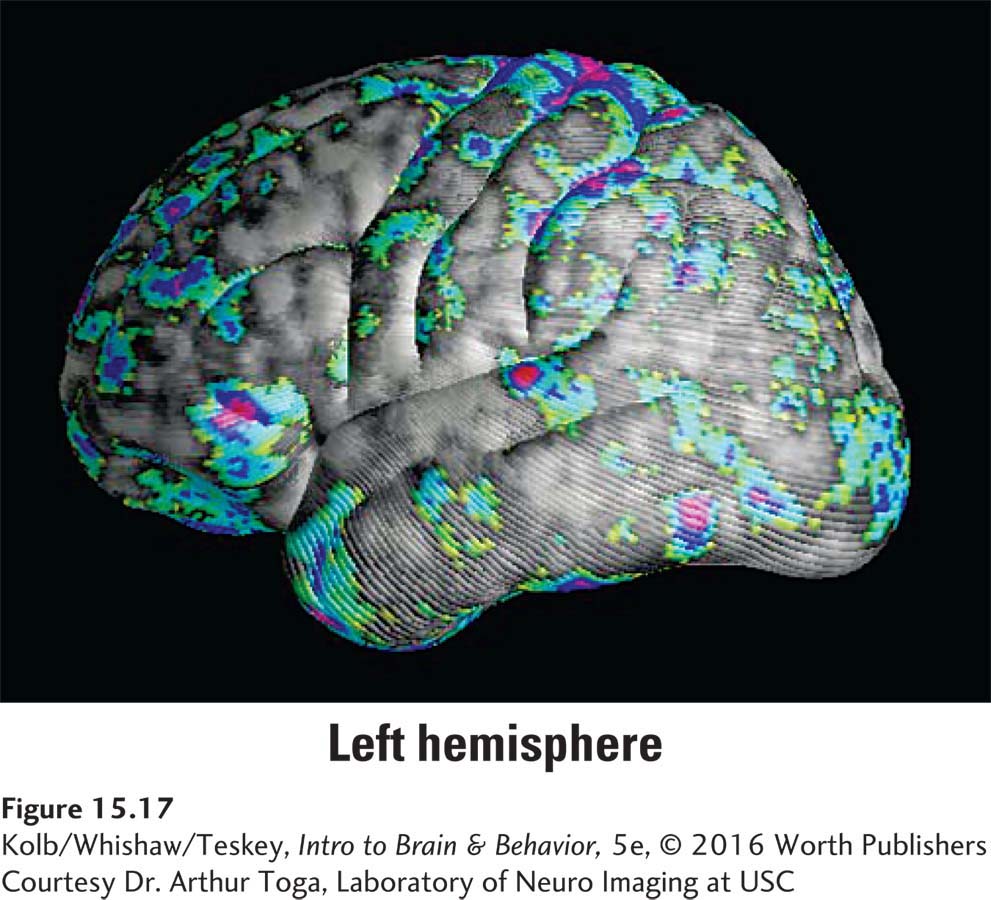
Another way to measure sex differences is cortical thickness independent of volume. Figure 15-17 shows that relative to men, women have increased cortical gray matter concentration in many cortical regions. Men’s gray matter concentration, by contrast, is more uniform across the cortex. The MRI studies represented in Figures 15-16 and 15-17 thus point to differences between men’s and women’s cortical organization.
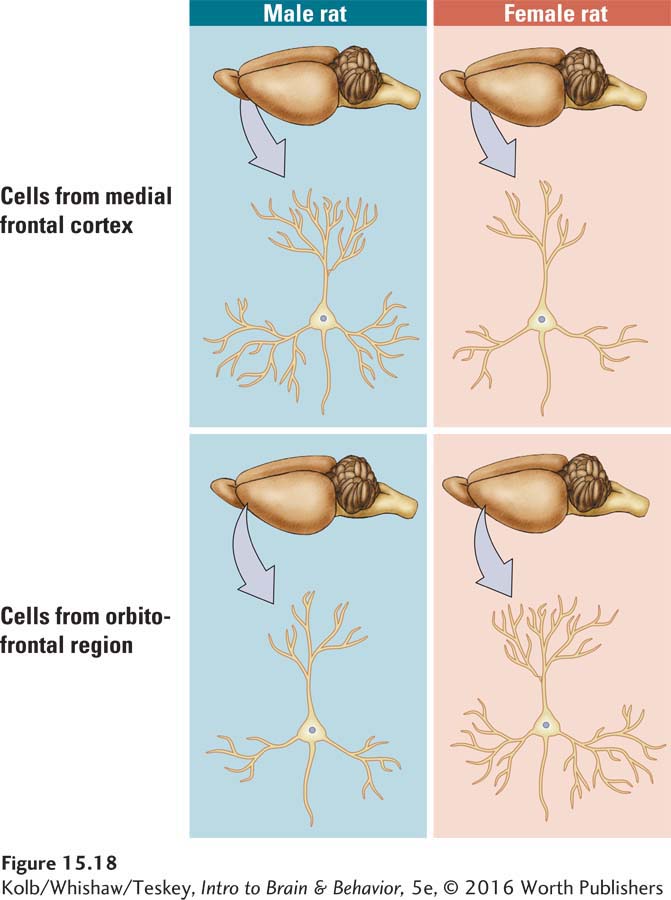
Sex differences in neuronal structure also exist. Gonadal hormones influence the structure of neurons in the prefrontal cortices of rats (Kolb & Stewart, 1991). The cells in one prefrontal region, located along the midline, have larger dendritic fields (and presumably more synapses) in males than in females, as shown in the top row of Figure 15-18 below. In contrast, the cells in the orbitofrontal region have larger dendritic fields (and presumably more synapses) in females than in males, as shown in the bottom row. These sex differences are not found in rats that have had their testes or ovaries removed at birth. Presumably, sex hormones somehow change the brain’s organization and ultimately its cognitive processing.
Stewart and Kolb (1994) found that the presence or absence of gonadal hormones affects the brain in adulthood as well as in early development. In this study, which focused on how hormones affect recovery from brain damage, middle-
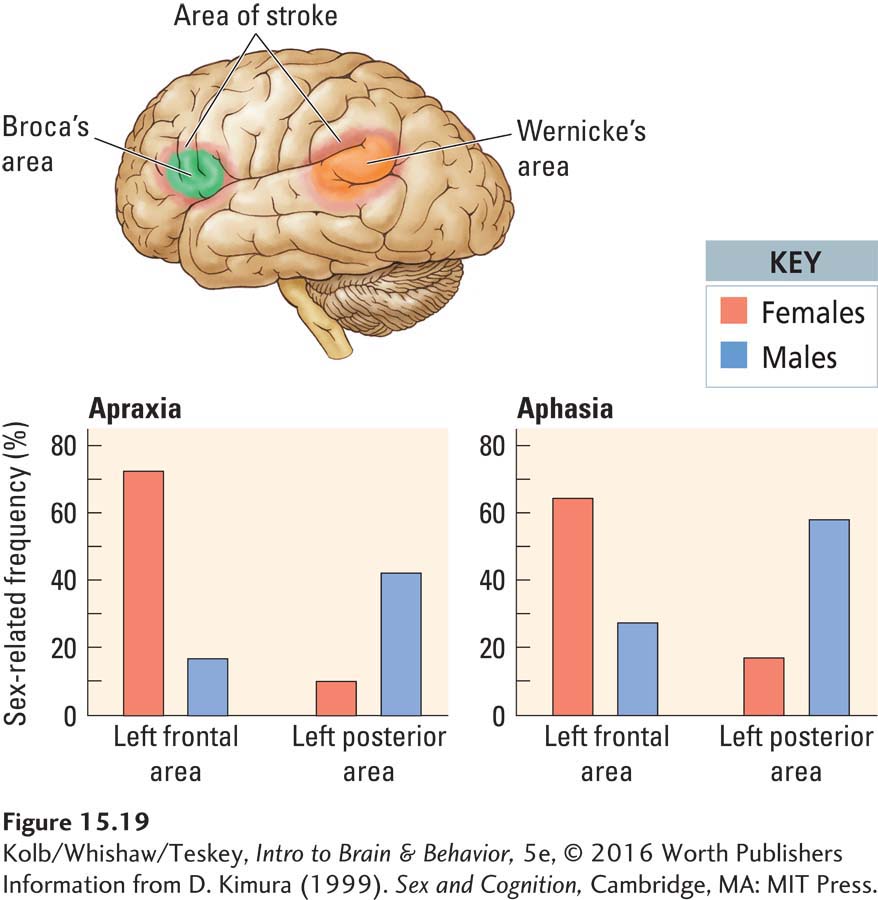
An additional way to consider the neural basis of sex differences is to look at the effects of cortical injury in men and women. If sex differences exist in the neural organization of cognitive processing, differences in the effects of cortical injury in the two sexes should appear. Doreen Kimura (1999) conducted this kind of study and showed that the pattern of cerebral organization within each hemisphere may in fact differ between the sexes.
548
Investigating people who had a cortical stroke in adulthood, Kimura tried to match the location and extent of injury in her male and female subjects. She found that men and women were almost equally likely to be aphasic subsequent to left-
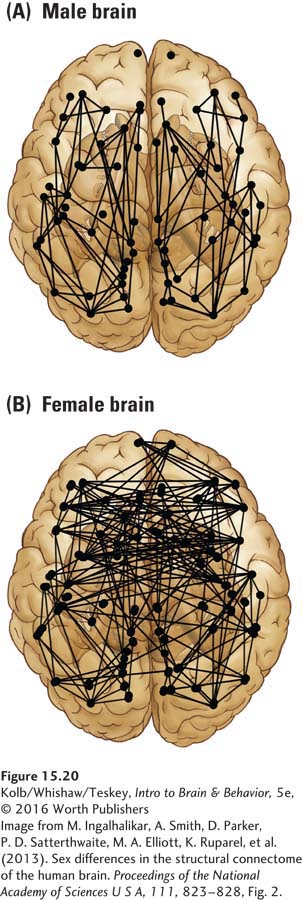
Section 12-5 describes the gender identity spectrum.
Evidence strongly favors a neural basis for gender identity, including transgender identity. Georg Krantz and colleagues (2014) used diffusion tensor analysis to compare brain networks of female-
That is, DTIs of the white matter microstructure of transgender females and males falls halfway between that of gender-
Evolution of Sex-
Although gonadal hormones have taken center stage in explaining sex differences in cognitive function, we are still left to question how these differences arose. To answer this question, we must look back at human evolution. Mothers pass their genes to both sons and daughters, and fathers do the same. Ultimately then, males and females of a species have virtually all their genes in common.
The only way a gene can affect one sex preferentially is for the animal’s gonadal hormones to influence the gene’s activities. Gonadal hormones are in turn determined by the presence or absence of the Y chromosome, which carries a gene called the testes-
Like other body organs, the brain is a potential target of natural selection. We should therefore expect to find sex-
549
Explaining sex-
Support for this hypothesis comes from comparing spatial problem solving in males of closely related mammalian species—
Meadow voles display far superior spatial skills to those of pine voles. A sex difference in spatial ability among meadow voles favors males, but pine voles have no such sex difference. The hippocampus is implicated in spatial navigation skills. Significantly, the hippocampus is larger in meadow voles than in pine voles, and it is larger in male meadow voles than in females (Gaulin, 1992). A similar logic could help explain sex-
Explaining sex-
Although such speculations are interesting, they are not testable. We will probably never know with certainty why sex-
Handedness and Cognitive Organization
550
Nearly everyone prefers one hand to the other for writing or throwing a ball. Most people prefer the right hand. In fact, left-
Because the left hemisphere controls the right hand, the general assumption is that right-
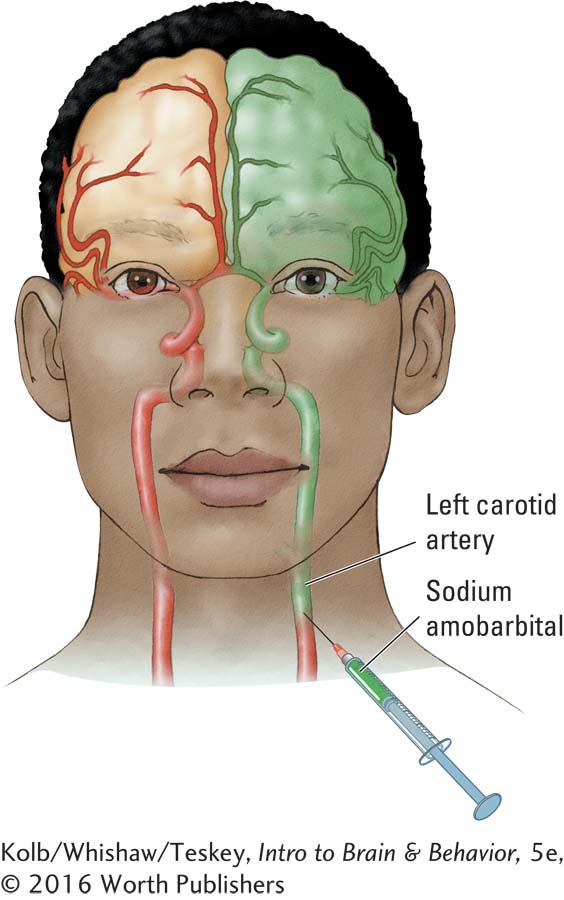
In the course of preparing patients with epilepsy for surgery to remove the abnormal tissue causing their seizures, Ted Rasmussen and Brenda Milner (1977) injected the left or right hemisphere with sodium amobarbital. This drug produces a short-
CLINICAL FOCUS 15-5
Sodium Amobarbital Test
Guy, a 32-
The complication with this surgery is that removing vessels sitting over the posterior speech zone poses a serious risk of permanent aphasia. Because Guy was left-
To achieve certainty in such doubtful cases, Jun Wada and Ted Rasmussen (1960) pioneered the technique of injecting sodium amobarbital, a barbiturate, into the carotid artery to briefly anesthetize the ipsilateral hemisphere. (Injections are now usually made through a catheter inserted into the femoral artery.) The procedure enables an unequivocal localization of speech because injection into the speech hemisphere results in speech arrest lasting as long as several minutes. As speech returns, it is characterized by aphasic errors.
Injection into the nonspeaking hemisphere may produce no or only brief speech arrest. The amobarbital procedure has the advantage of allowing each hemisphere to be studied separately in the functional absence of the other (anesthetized) hemisphere. Because the period of anesthesia lasts several minutes, a variety of functions, including memory and movement, can be studied to determine a hemisphere’s capabilities.
The sodium amobarbital test is always performed bilaterally, with the second cerebral hemisphere being injected several days after the first one to make sure that no residual drug effect lingers. In the brief period of drug action, the patient is given a series of simple tasks requiring the use of language, memory, and object recognition. To test speech, the patient is asked to name some common objects presented in quick succession, to count, to recite the days of the week forward and backward, and to spell simple words.
If the injected hemisphere is nondominant for speech, the patient may continue to carry out the verbal tasks, although there is often a period as long as 30 seconds during which he or she appears confused and is silent but can resume speech with urging. When the injected hemisphere is dominant for speech, the patient typically stops talking and remains completely aphasic until recovery from the anesthesia is well along, somewhere in the range of 4 to 10 minutes.
Guy was found to have speech in the left hemisphere. During the test of his left hemisphere, he could not talk. Later, he said that when he was asked about a particular object, he wondered just what the question meant. When he finally had some vague idea, he had no idea of what the answer was or how to say anything. By then he realized that he had been asked all sorts of other questions to which he had also not responded.
When asked which objects he had been shown, he said he had no idea. However, when given an array of objects and asked to choose with his left hand, he identified the objects by pointing because his nonspeaking right hemisphere controlled that hand. In contrast, his speaking left hemisphere had no memory of the objects: it had been asleep.
Rasmussen and Milner found that in virtually all right-
551
Sandra Witelson and Charlie Goldsmith (1991) asked whether any other gross differences in the brain structure of right-
Whether this enlarged callosum is due to a greater number of fibers, to thicker fibers, or to more myelin remains to be seen. If the larger corpus callosum is due to more fibers, the difference would be on the order of 25 million more fibers. Presumably, such a difference would have major implications for the organization of cognitive processing in left-
Synesthesia
Some variations in brain organization are idiosyncratic rather than systematic. Synesthesia is an individual’s capacity to join sensory experiences across sensory modalities, as discussed in Clinical Focus 15-6, A Case of Synesthesia. Examples include the ability to hear colors or taste shapes. Edward Hubbard (2007) estimated the incidence of synesthesia at about 1 in 23 people, although for most it likely is limited in scope.
CLINICAL FOCUS 15-6
A Case of Synesthesia
Michael Watson tastes shapes. His sensory joining first came to the attention of neurologist Richard Cytowic over dinner. After tasting a sauce he was making for roast chicken, Watson blurted out, “There aren’t enough points on the chicken.”
When Cytowic quizzed him about this strange remark, Watson said that all flavors had shape for him. “I wanted the taste of this chicken to be a pointed shape, but it came out all round. Well, I mean it’s nearly spherical. I can’t serve this if it doesn’t have points” (Cytowic, 1998, p. 4).
Watson has synesthesia, which literally means feeling together. All his life Watson has experienced the feeling of shape when he tastes or smells food. When he tastes intense flavors, he reports an experience of shape that sweeps down his arms to his fingertips. He experiences the feeling of weight, texture, warmth or cold, and shape, just as though he were grasping something.
The feelings are not confined to his hands, however. Watson experiences some taste shapes, such as points, over his whole body. He experiences others only on the face, back, or shoulders. These impressions are not metaphors, as other people might use when they say that a cheese is sharp or a wine is textured. Such descriptions make no sense to Watson. He actually feels the shapes.
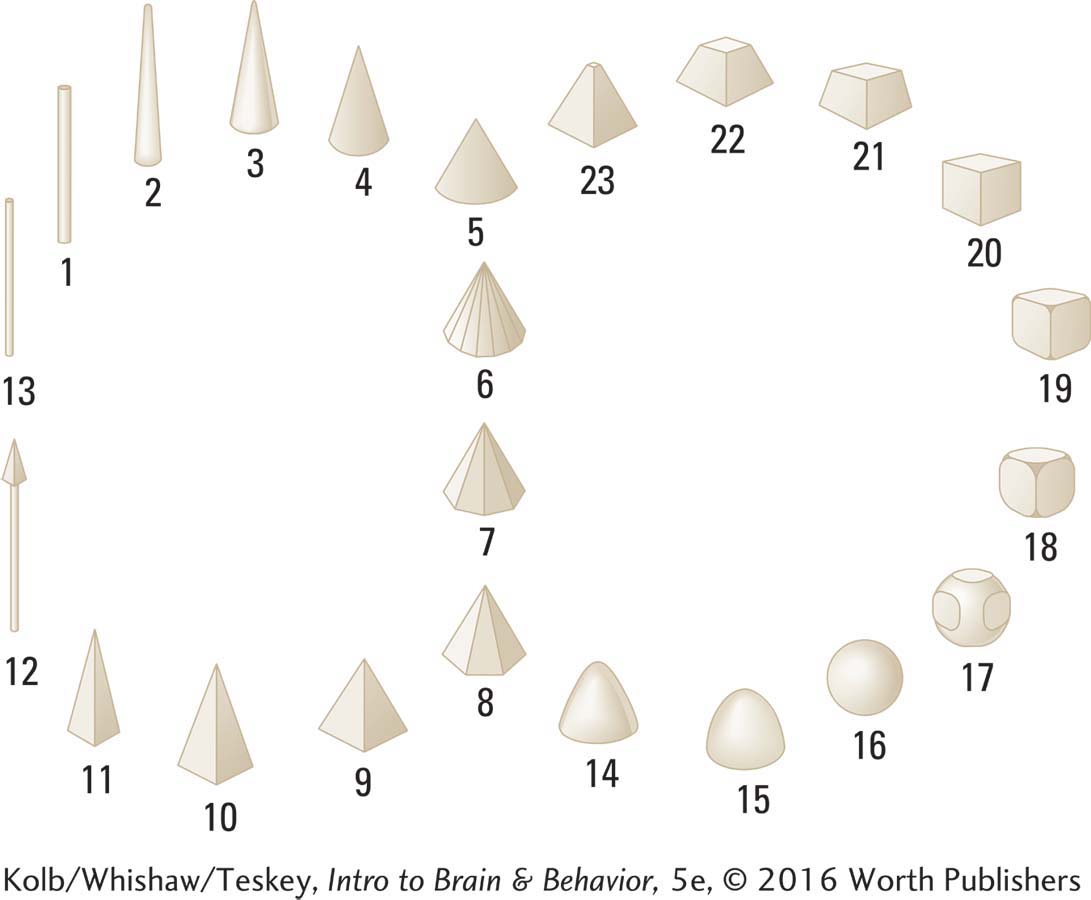
Cytowic systematically studied Watson to determine whether his feelings of shape were always associated with particular flavors and found that they were. Cytowic devised the set of geometric figures shown here to allow Watson to communicate which shapes he associated with various flavors.
Musician–
Synesthesia runs in families—
552
If you’ve shivered on hearing a particular piece of music or the noise of fingernails scratching across a chalkboard, you have felt sound. Even so, other sensory blendings may be difficult to imagine. How can sounds or letters possibly produce colors? Studies of synesthetes show that the same stimuli always elicit the same experiences for them.
The most common form of synesthesia is colored hearing. For many synesthetes, this means hearing both speech and music in color—
The five primary senses (vision, hearing, touch, taste, and smell) all generate synesthetic pairings. Most, however, are in one direction. For instance, whereas synesthetes may see colors when they hear, they do not hear sounds in colors. Furthermore, some sensory combinations occur rarely, if at all. In particular, taste or smell rarely triggers a synesthetic response like Michael Watson’s.
Because each case is idiosyncratic, synesthesia’s neurological basis is difficult to investigate. Few studies have related it directly to brain function or brain organization, and different people may experience it for different reasons. Various hypotheses have been advanced to account for synesthesia:
Extraordinary neural connections between the sensory regions are related in a particular synesthete.
Activity is increased in the frontal lobe multimodal cortex, which receives inputs from more than one sensory area.
Particular sensory inputs elicit unusual patterns of cerebral activation.
Whatever the explanation, when it comes to certain sensory inputs, the brain of a synesthete certainly works differently from other people’s brains.
15-5 REVIEW
Variations in Cognitive Organization
Before you continue, check your understanding.
Question 1
The two major contributors to organizational differences in individual brains are ____________ and ____________.
Question 2
Differences in the cerebral organization of thinking are probably related to differences in the ____________ that underlie different types of cognitive processing.
Question 3
People who experience certain sensations in more than one sensory modality are said to have ____________.
Question 4
What roles do gonadal hormones play in brain organization and function?
Answers appear in the Self Test section of the book.