The endomembrane system is a group of interrelated organelles
Much of the volume of some eukaryotic cells is taken up by an extensive endomembrane system. This is an interconnected system of membrane-
focus: key figure
Question
Q: What processes in the endomembrane system are mediated by vesicles?
Assembly of the ER from the nuclear envelope; transport between Golgi apparatus cisternae; transport from ER to Golgi apparatus; endocytosis; exocytosis; transport from Golgi apparatus to cell membrane.
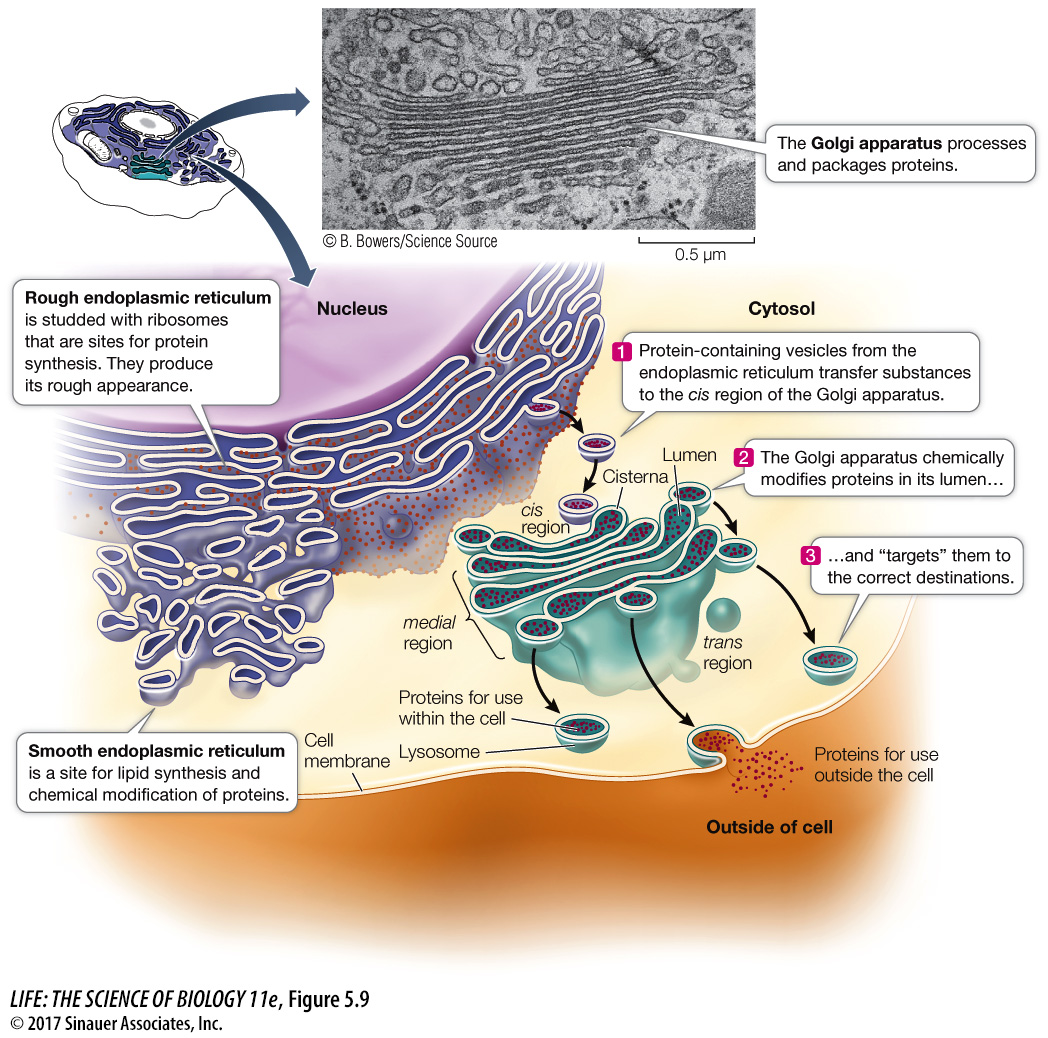
ENDOPLASMIC RETICULUM Electron micrographs of eukaryotic cells reveal networks of interconnected membranes branching throughout the cytoplasm, forming tubes and flattened sacs. These membranes are collectively called the endoplasmic reticulum, or ER. The interior compartment of the ER, referred to as the lumen, is separate and distinct from the surrounding cytoplasm (see Figure 5.9). The ER can enclose up to 10 percent of the interior volume of the cell, and its folds result in a surface area many times greater than that of the cell membrane. There are two types of endoplasmic reticulum, the so-
 Rough endoplasmic reticulum (RER) is called “rough” because of the many ribosomes attached to the outer surface of the membrane, giving it a “rough” appearance in electron microscopy (see Figure 5.7). The bound ribosomes are actively involved in protein synthesis, but that is not the entire story:
Rough endoplasmic reticulum (RER) is called “rough” because of the many ribosomes attached to the outer surface of the membrane, giving it a “rough” appearance in electron microscopy (see Figure 5.7). The bound ribosomes are actively involved in protein synthesis, but that is not the entire story:
The RER receives into its lumen certain newly synthesized proteins (including proteins destined for lysosomes, the cell membrane, and export from the cell), segregating them away from the cytoplasm. The RER also participates in transporting these proteins to other locations in the cell.
While inside the RER, proteins can be chemically modified to alter their functions and to “tag” them for delivery to specific cellular destinations.
Proteins are shipped to destinations elsewhere in the cell enclosed within vesicles that pinch off from the RER.
Most membrane-
bound proteins are made in the RER.
A protein enters the lumen of the RER through a pore as it is synthesized. As with a protein passing through a nuclear pore, there is a special transport mechanism to get proteins into the ER, which you’ll learn about in Chapter 14. Once in the lumen of the RER, proteins undergo several changes, including the formation of disulfide bridges and folding into their tertiary structures (see Figure 3.5).
Some proteins are covalently linked to carbohydrate groups in the RER, thus becoming glycoproteins. In the case of proteins directed to the lysosomes, the carbohydrate groups are part of an “addressing” system that ensures that the right proteins are directed to those organelles. This addressing system is very important because the enzymes within the lysosomes are some of the most destructive the cell makes. Were they not properly addressed and contained, they could destroy the cell.
93
 The smooth endoplasmic reticulum (SER) lacks ribosomes and is more tubular (and less like flattened sacs) than the RER, but it shows continuity with portions of the RER (see Figure 5.9). Certain proteins that are synthesized in the RER are chemically modified within the lumen of the *SER. The SER has four other important roles:
The smooth endoplasmic reticulum (SER) lacks ribosomes and is more tubular (and less like flattened sacs) than the RER, but it shows continuity with portions of the RER (see Figure 5.9). Certain proteins that are synthesized in the RER are chemically modified within the lumen of the *SER. The SER has four other important roles:
It is responsible for the chemical modification of small molecules taken in by the cell that may be toxic to the cell. These modifications make the targeted molecules more polar, so they are more water-
soluble and easily removed. It is the site for glycogen degradation in animal cells. We will discuss this important process in Chapter 9.
It is the site where lipids and steroids are synthesized, as well as some polysaccharides in plant cells.
It stores calcium ions, which when released trigger a number of cell responses.
*connect the concepts Regulating muscle contraction is one of the important roles played by the calcium ions stored by the SER. See Key Concept 47.1.
94
Cells that synthesize a lot of protein for export are usually packed with RER. Examples include glandular cells that secrete digestive enzymes and white blood cells that secrete antibodies. In contrast, cells that carry out less protein synthesis (such as storage cells) contain less RER. Liver cells, which modify molecules (including toxins) that enter the body from the digestive system, have abundant SER.
 GOLGI APPARATUS The Golgi apparatus (or Golgi complex, or just Golgi) is another part of the diverse, dynamic, and extensive endomembrane system (see Figure 5.9). This structure was named after its discoverer, Camillo Golgi. Its appearance varies, but it almost always consists of two components: flattened membranous sacs called cisternae (singular cisterna) that are piled up like saucers, and small membrane-
GOLGI APPARATUS The Golgi apparatus (or Golgi complex, or just Golgi) is another part of the diverse, dynamic, and extensive endomembrane system (see Figure 5.9). This structure was named after its discoverer, Camillo Golgi. Its appearance varies, but it almost always consists of two components: flattened membranous sacs called cisternae (singular cisterna) that are piled up like saucers, and small membrane-
Animation 5.1 The Golgi Apparatus
www.life11e.com/
The Golgi apparatus has several roles:
It receives protein-
containing vesicles from the RER. It modifies, concentrates, packages, and sorts proteins before they are sent to their cellular or extracellular destinations.
It adds carbohydrates to proteins and modifies other carbohydrates that were attached to proteins in the RER.
It is where some polysaccharides for the plant cell wall are synthesized.
The cisternae of the Golgi apparatus have three functionally distinct regions:
The cis region lies nearest to the nucleus or a patch of RER.
The trans region lies closest to the cell membrane.
The medial region lies in between (see Figure 5.9).
The terms cis, trans, and medial derive from Latin words meaning, respectively, “on the same side,” “on the opposite side,” and “in the middle.” These three parts of the Golgi apparatus contain different enzymes and perform different functions.
Protein-
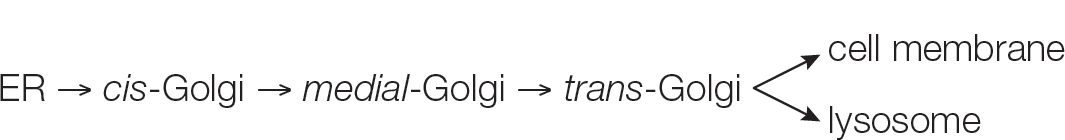
How do the vesicles in this system recognize their targets for fusion? In other words, when a vesicle buds off from the ER, how does it “know” to fuse with the cis-Golgi membrane? The answer lies in proteins on both membranes appropriately called SNARES. Pairs of SNARES are complementary—
LYSOSOMES The primary lysosomes originate from the Golgi apparatus. They contain digestive enzymes and are the sites where macromolecules—
R1—R2 (linked monomers) + H2O → R1—OH + R2—H
Lysosomes are about 1µm in diameter, are surrounded by a single membrane, and have a densely staining, featureless interior (Figure 5.10). There may be dozens of lysosomes in a cell, depending on its needs.
Activity 5.4 Lysosomal Digestion
www.life11e.com/
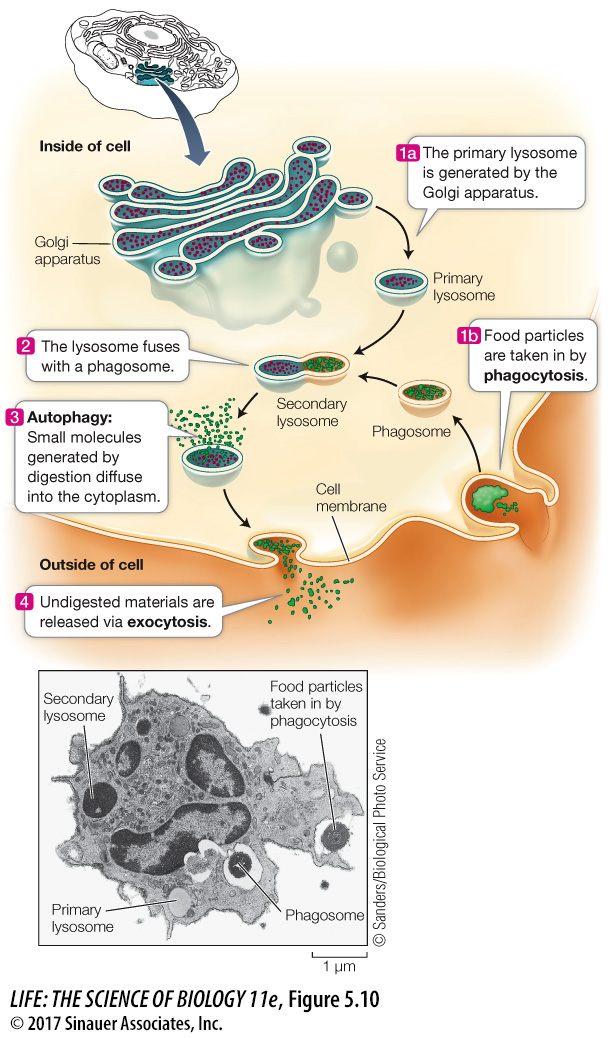
Lysosomes are cellular “disposals,” compartments for the breakdown of food, other cells, or foreign objects that are taken up by the cell. These materials enter the cell by a process called phagocytosis (phago, “eat,” + cytosis, “cellular”). In this process, a pocket forms in the cell membrane and then deepens and encloses material from outside the cell. The pocket becomes a small vesicle called a phagosome, containing food or other material, which breaks free of the cell membrane to move into the cytoplasm. The phagosome fuses with a primary lysosome to form a secondary lysosome, where digestion occurs.
The effect of this fusion is rather like children devouring Halloween candy: the enzymes in the secondary lysosome quickly hydrolyze the food particles. These reactions are enhanced by the acidity of the lysosome’s interior, where the pH is lower than in the surrounding cytoplasm. The products of digestion pass through the membrane of the lysosome, providing energy and raw materials for other cellular processes. For example, a protease might hydrolyze proteins to amino acids, which are released to the cytoplasm for reassembly into other proteins. The “used” secondary lysosome, now containing undigested particles, then moves to the cell membrane, fuses with it, and releases the undigested contents to the environment, a process called exocytosis.
Phagocytes (see Key Concept 41.1) are specialized cells that have an essential role in taking up and breaking down materials; they are found in nearly all animals and many protists. You will encounter them and their activities again at many places in this book, but at this point one example suffices: in the human liver and spleen, phagocytes digest approximately 10 billion aged or damaged blood cells each day! The digestion products are then used to make new cells to replace those that are digested.
Lysosomes are active even in cells that do not perform phagocytosis. Cells are dynamic systems; some cell components are continually being broken down and replaced by new ones. The programmed destruction of cell components is called autophagy, and lysosomes are where the cell breaks down its own materials. With the proper signal, lysosomes can engulf entire organelles, hydrolyzing their constituents.
95
How important is autophagy? An entire class of human diseases called lysosomal storage diseases are caused by the failure of lysosomes to digest specific cellular components; these diseases are invariably harmful or fatal. An example is Tay-
Plant cells do not appear to contain lysosomes, but the vacuole of a plant cell may function in an equivalent capacity because it, like lysosomes, contains many digestive enzymes.