Linkage is revealed by studies of the sex chromosomes
In Mendel’s work, reciprocal crosses always gave similar results; it did not matter whether a dominant allele was contributed by the female parent or the male parent. But in some cases, the parental origin of a chromosome does matter. For example, human males inherit a bleeding disorder called hemophilia from their mother, not from their father. To understand the types of inheritance in which the parental origin of an allele is important, we must consider the ways in which sex is determined in different species.
SEX DETERMINATION BY CHROMOSOMES In corn, every diploid adult has both male and female reproductive structures. The tissues in these two types of structure are genetically identical, just as roots and leaves are genetically identical. Organisms such as corn, in which the same individual produces both male and female gametes, are said to be monoecious (Greek, “one house”). Other organisms, such as date palms and most animals, are dioecious (“two houses”), meaning that some individuals can produce only male gametes and others can produce only female gametes. In other words, in dioecious organisms the different sexes are different individuals.
In mammals and birds, sex is determined by differences in the chromosomes, but such determination operates in different ways in different groups of organisms. For example, in many animals, including mammals, sex is determined by a single pair of sex chromosomes, which differ from one another. The remaining chromosomes, called autosomes, occur in pairs in males and females. For example, in humans there are 22 pairs of autosomes in males and females, and 1 pair of sex chromosomes. The chromosomal bases for sex determination in various groups of animals are summarized in Table 12.2.
| Animal group | Mechanism |
|---|---|
| Bees | Males are haploid, females are diploid |
| Fruit flies | Fly is female if ratio of X chromosomes to sets of autosomes is 1 or more |
| Birds | Males ZZ (homogametic), females WZ (heterogametic) |
| Mammals | Males XY (heterogametic), females XX (homogametic) |
259
The sex chromosomes of female mammals consist of a pair of X chromosomes. Male mammals, by contrast, have one X chromosome and a sex chromosome that is not found in females, the Y chromosome. Females may be represented as XX (homogametic) and males as XY (heterogametic).
MALE MAMMALS PRODUCE TWO KINDS OF GAMETES Each gamete produced by a male mammal has a complete set of autosomes, but just as each gamete carries one copy of each pair of autosomes, half carry a Y. When an X-
SEX CHROMOSOME ABNORMALITIES REVEALED THE GENE THAT DETERMINES SEX Can we determine which chromosome, X or Y, carries the sex-
We can learn something about the functions of X and Y chromosomes from abnormal sex chromosome arrangements resulting from nondisjunction during meiosis or mitosis (see Key Concept 11.5). As you will recall, nondisjunction occurs when a pair of homologous chromosomes (in meiosis I) or sister chromatids (in mitosis or meiosis II) fails to separate. As a result, a gamete may have one too few or one too many chromosomes. If this gamete fuses with another gamete that has the full haploid chromosome set, the resulting offspring will be aneuploid, with fewer or more chromosomes than normal.
In humans, XO individuals sometimes appear. The O indicates that a chromosome is missing—
These observations suggested that the gene controlling maleness is located on the Y chromosome. Observations of people with other types of chromosomal abnormalities helped researchers pinpoint the location of that gene:
Some women are genetically XY but lack a small portion of the Y chromosome.
Some men are genetically XX but have a small piece of the Y chromosome attached to another chromosome.
The Y fragments that are respectively missing and present in these two cases are the same and contain the maleness-
The SRY gene encodes a protein involved in primary sex determination—that is, the determination of the kinds of gametes that an individual will produce and the organs that will make them (the male and female gonads). In the presence of the functional SRY protein, an embryo develops sperm-
One function of the gonads is to produce hormones (such as testosterone and estrogen) that send signals to the rest of the body and control the development of secondary sex characteristics. These are outward manifestations of maleness and femaleness, such as differences in body type, breast development, body hair, and voice. Secondary sex characteristics distinguish males and females but are not directly part of the reproductive system.
SEX-
Thomas Morgan identified a sex-
Morgan crossed a homozygous red-
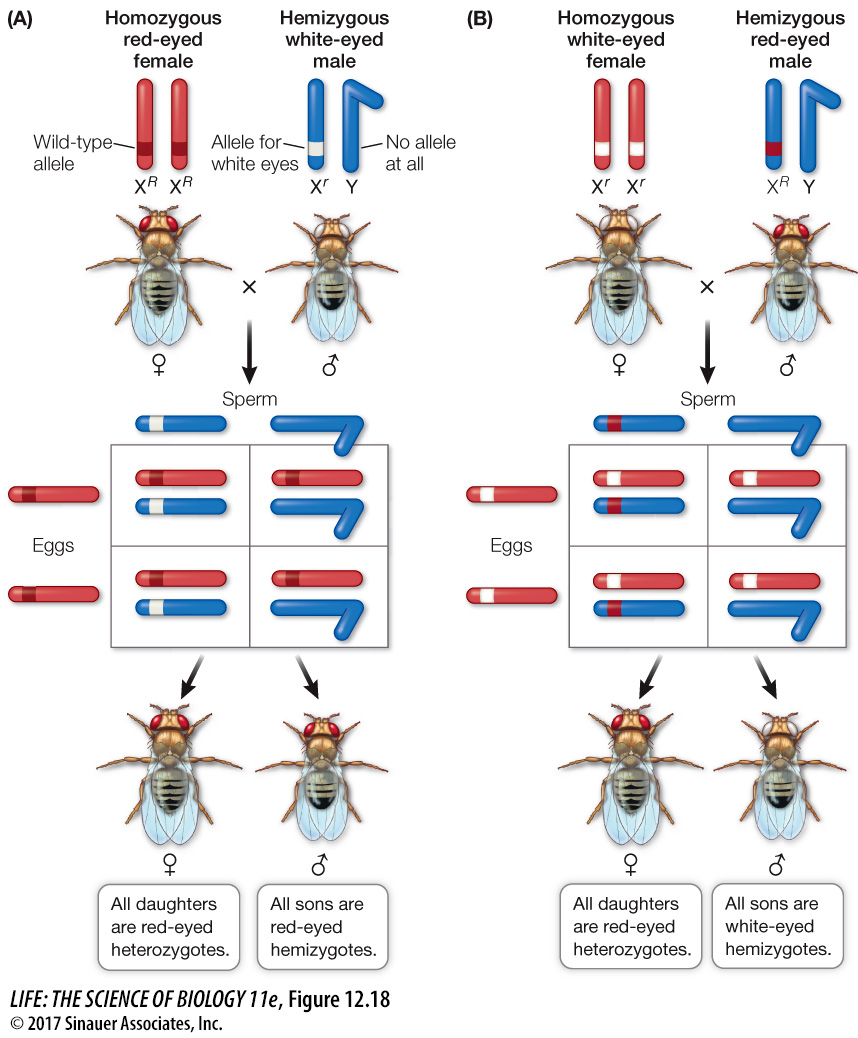
Question
Q: If a red-
Heterozygous, XRXr. The female parent passed the Xr chromosome to the offspring, which had a Y chromosome from the male parent.
When Morgan performed the reciprocal cross, in which a white-
260
These and other experiments led to the term sex-
Many sexually reproducing species, including humans, have sex chromosomes. As in most fruit flies, human males are XY, females are XX, and relatively few of the genes that are present on the X chromosome are present on the Y. Pedigree analyses of X-
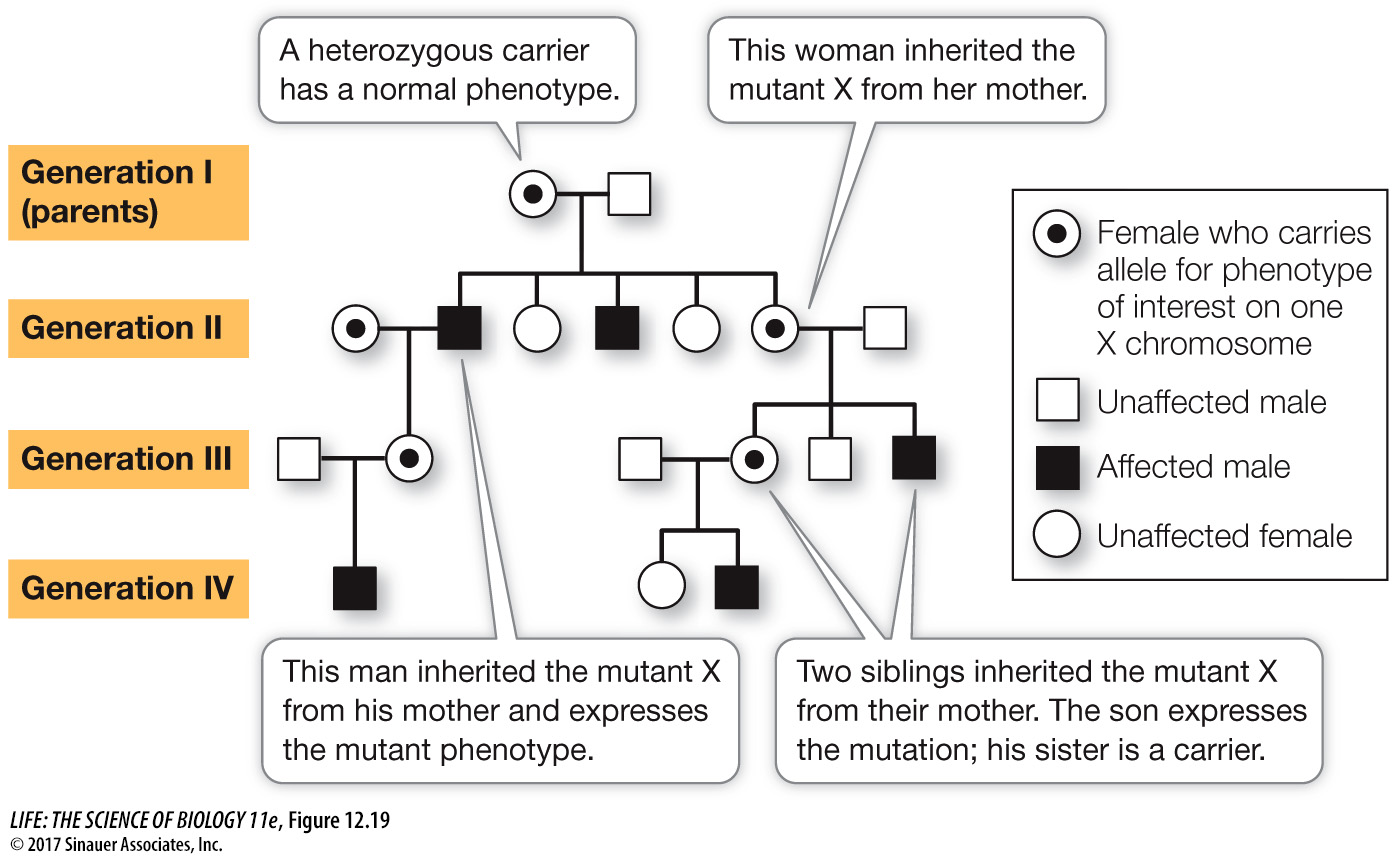
The phenotype appears much more often in males than in females, because only one copy of the rare allele is needed for its expression in males, whereas two copies must be present in females.
A male with the mutation can pass it on only to his daughters; all his sons get his Y chromosome.
Daughters who receive one X-
linked mutation are heterozygous carriers. They are phenotypically normal, but they can pass the mutant allele to their sons or daughters. On average, half their children will inherit the mutant allele since half of their X chromosomes carry the normal allele. The mutant phenotype can skip a generation if the mutation passes from a male to his daughter (who will be phenotypically normal) and then to her son.