Nonrandom mating can change genotype or allele frequencies
Mating patterns often alter genotype frequencies because the individuals in a population do not choose mates at random. For example, self-
433
Many nonrandom mating systems produce changes in genotype frequencies but not in allele frequencies, and thus do not, by themselves, result in evolutionary change in a population. However, nonrandom mating systems that result in different reproductive success among individuals do produce allele frequency changes from one generation to the next. One such process, sexual selection occurs when individuals of one sex mate preferentially with particular individuals of the opposite sex rather than at random.
Sexual selection was first suggested by Charles Darwin, who developed the idea to explain the evolution of conspicuous traits that would appear to inhibit survival, such as bright colors and elaborate courtship displays in males of many species. He hypothesized that these features either improved the ability of their bearers to compete for access to mates (intrasexual selection) or made their bearers more attractive to members of the opposite sex (intersexual selection).
Darwin argued that while natural selection typically favors traits that enhance the survival of their bearers or their bearers’ descendants, sexual selection is primarily about successful reproduction. An animal that survives but fails to reproduce makes no contribution to the next generation. Thus sexual selection may favor traits that enhance an individual’s chances of reproduction even when these traits reduce its chances of survival. For example, females may be more likely to see or hear males with a given trait (and thus be more likely to mate with those males), even though the favored trait also increases the chances that the male will be seen or heard by a predator.
One example of a trait that Darwin attributed to sexual selection is the remarkable tail of the male African long-

experiment
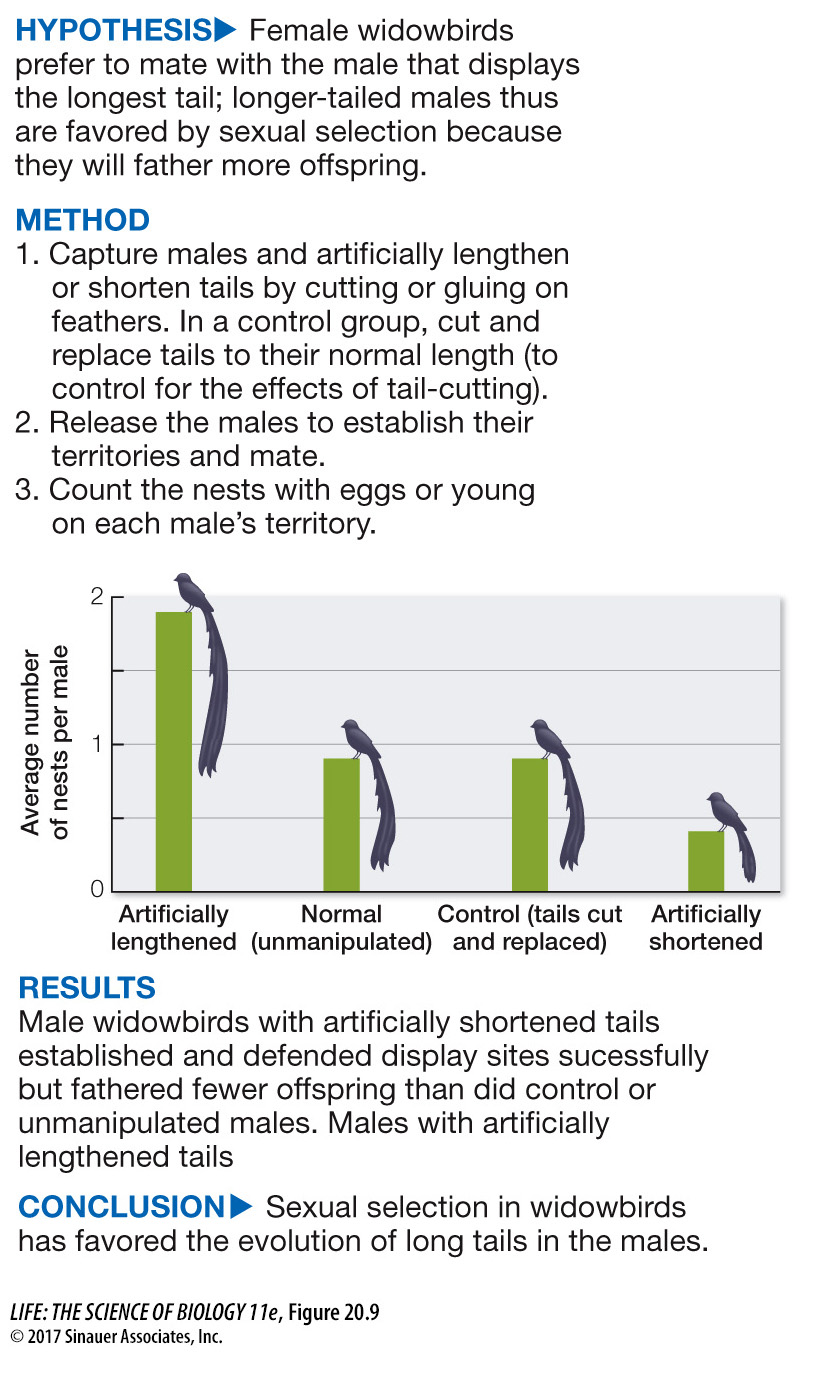
Figure 20.9 Sexual Selection in Action
Original Paper: Andersson, M. 1982. Female choice selects for extreme tail length in a widowbird. Nature 299: 818–
Behavioral ecologist Malte Andersson tested Darwin’s hypothesis that excessively long tails evolved in male widowbirds because female preference for longer-