Heterozygote advantage maintains polymorphic loci
In many cases, different alleles of a particular gene are advantageous under different environmental conditions. Most organisms experience a wide variety of environmental conditions over time. A night is dramatically different from the preceding day. A cold, cloudy day differs from a clear, hot one. Day length and temperature change seasonally. For many genes, a single allele is unlikely to perform well under all these conditions. In such cases, heterozygous individuals (with two different alleles) are likely to outperform individuals that are homozygous (with only one of those two alleles).
Colias butterflies of the Rocky Mountains live in environments where dawn temperatures are often too cold, and afternoon temperatures too hot, for the butterflies to fly. Populations of these butterflies are polymorphic for a gene that encodes the enzyme phosphoglucose isomerase (PGI), which influences how well a butterfly flies at different temperatures. Butterflies with certain PGI genotypes can fly better during the cold hours of early morning; others perform better during midday heat (Figure 20.17). The optimal body temperature for flight is 35°C–
441
experiment
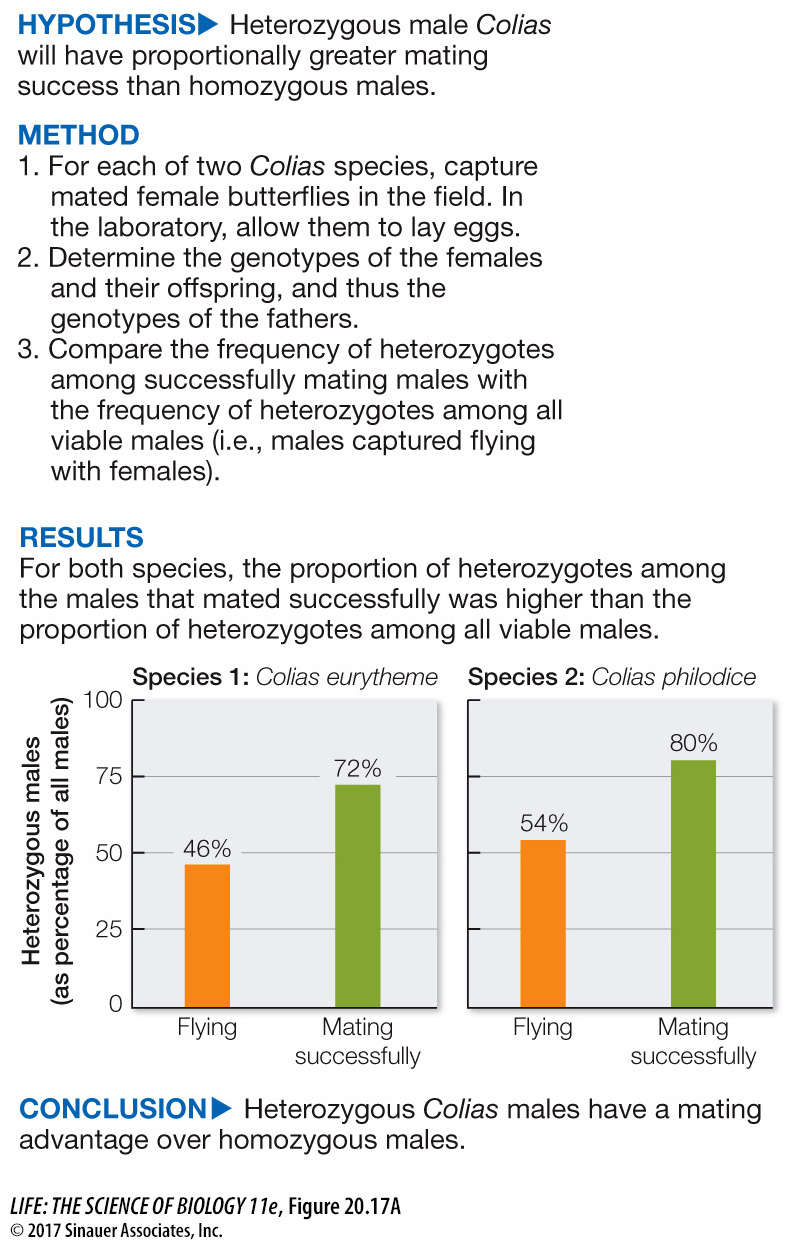
Figure 20.17A A Heterozygote Mating Advantage
Original Paper: Watt, W. B., P. A. Carter and S. M. Blower. 1985. Adaptation at specific loci. IV. Differential mating success among glycolytic allozyme genotypes of Colias butterflies. Genetics 109: 157–
Among butterflies of the genus Colias, males that are heterozygous for two alleles of the PGI enzyme can fly farther under a broader range of temperatures than males that are homozygous for either allele. Does this ability give heterozygous males a mating advantage?
work with the data
Figure 20.17B Do Heterozygous Males Have a Mating Advantage?
Original Paper: Watt, W. B., P. A. Carter and S. M. Blower. 1985.
Ward Watt and his colleagues tested the hypothesis that males with two different alleles for the PGI enzyme (heterozygotes) were more likely to mate successfully with females than were homozygous males. They reasoned that the heterozygous males could fly farther under a broader range of temperatures than could homozygous males, and that this ability would give heterozygous males greater access to receptive females. To test this hypothesis, they needed to know the frequency of heterozygotes among successfully mating males, and they needed to compare that frequency with the frequency of heterozygotes among males in the general population (i.e., all the potential mates available to females). To estimate the frequency of heterozygotes among mating males, Watt and his colleagues collected mated female butterflies in the field and allowed them to lay eggs in the laboratory. They hatched the eggs and determined the genotypes of the offspring, as well as the genotypes of the females. Using this information, they could determine the genotypes of the males that fathered the larvae. They then compared the estimated frequency of heterozygotes among the successful fathers with the frequency of heterozygotes among all viable males in the population. Samples of their data are given in the table.
| All viable malesa | Mating males | |||
|---|---|---|---|---|
| Species | Heterozygous/ total |
Percent heterozygous |
Heterozygous/ total |
Percent heterozygous |
| C. philodice | 32/74 | 43.2 | 31/50 | 62 |
| C. eurytheme | 44/92 | 47.8 | 45/59 | 76.3 |
|
a “Viable males” are males captured flying with females (hence the potential to mate). |
||||
QUESTIONS
Question 1
If we assume that the proportions of each genotype among mating males should be the same as the proportions seen among all viable males, what is the number of mating males expected to be heterozygous and homozygous in each sample?
For C. philodice, 43.2% of all viable males are heterozygous, so 56.8% must be homozygous. To get expected numbers of heterozygous and homozygous mating males, we multiple the expected proportions (from all viable males) by the total number of mating males sampled. Therefore we expect to see (0.432)(50) = 21.6 heterozygous mating males, and (0.568)(50) = 28.4 homozygous mating males. If we repeat the same calculations for C. eurytheme, we expect (under the given assumption) to see (0.478)(59) = 28.2 heterozygous mating males and (0.522)(59) = 30.8 homozygous mating males.
Question 2
Use a chi-
Chi-
| Genotype | Expected (E) | Observed (O) | O – E | (O – E)2 | (O – E)2/E |
|---|---|---|---|---|---|
| Heterozygotes | 21.6 | 31 | 9.4 | 88.36 | 4.091 |
| Homozygotes | 28.4 | 19 | –9.4 | 88.36 | 3.111 |
The sum of the last column gives the chi-
Chi-
| Genotype | Expected (E) | Observed (O) | O – E | (O – E)2 | (O – E)2/E |
|---|---|---|---|---|---|
| Heterozygotes | 28.2 | 45 | 16.8 | 282.17 | 10.005 |
| Homozygotes | 30.8 | 14 | –16.8 | 282.17 | 9.162 |
The sum of the last column gives the chi-
Question 3
The investigators determined the genotypes of enough larvae from each batch of eggs to judge the genotype of the father with 99% certainty. How many larvae did they need to measure to achieve that level of certainty?
Hint: If the female is homozygous—
(0.75)16 is approximately 0.01002. Therefore the investigators would need to analyze at least 16 larvae from each batch of eggs to judge the genotype of the father with 99 percent certainty. An easy way to find this answer is to multiply 0.75 × 0.75, and then multiply the answer by 0.75, and continue until the result is approximately 0.01. Keep track of the number of times you multiply by 0.75 to find the appropriate sample size.
A similar work with the data exercise may be assigned in LaunchPad.
442
Heterozygous Colias butterflies can fly over a greater range of temperatures than homozygous individuals, which should give them an advantage in foraging and finding mates. A test of this prediction found that heterozygous males did indeed have a mating advantage, and further, that this advantage maintains the polymorphism in the population. Of course, the heterozygous genotype can never become fixed in the population, because the offspring of two heterozygotes will always include both classes of homozygotes in addition to heterozygotes.