Much of evolution is neutral
In 1968, Motoo Kimura proposed the neutral theory of molecular evolution. Kimura suggested that, at the molecular level, the majority of the variants we observe in most populations are selectively neutral; that is, they confer neither an advantage nor a disadvantage on their bearers. These neutral variants accumulate through genetic drift rather than through positive selection.
The rate of fixation of neutral mutations by genetic drift is independent of population size. To see why this is so, consider a diploid population of size N and a neutral mutation rate μ (mu) per gamete per generation at a locus. The number of new mutations would be, on average, μ × 2N, because 2N gene copies are available to mutate. The probability that a given mutation will be fixed by drift alone is its frequency, which equals 1/(2N) for a newly arisen mutation. We can multiply these two terms to get the rate of fixation of neutral mutations (m) in a given population of N individuals:
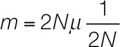
Therefore the rate of fixation m of neutral mutations depends only on the neutral mutation rate μ and is independent of population size. A given mutation is more likely to appear in a large population than in a small one, but any mutation that does appear is more likely to become fixed in a small population. These two influences of population size cancel each other out, so the rate of fixation of neutral mutations is equal to the mutation rate (i.e., m = μ).
As long as the underlying mutation rate is constant, macromolecules evolving in separate populations should diverge from one another in neutral changes at a constant rate. Investigators have confirmed that the rate of evolution of particular genes and proteins is often relatively constant over time and can therefore can be used as a “molecular clock.” As we described in Key Concept 21.3, molecular clocks can be used to calculate evolutionary divergence times between species.
Although much of the genetic variation we observe in populations is the result of neutral evolution, the neutral theory does not imply that most mutations have no effect on the organism. Many mutations are never observed in populations because they are lethal or strongly detrimental to the organism and are thus quickly removed from the population through natural selection. Similarly, mutations that confer a selective advantage tend to be quickly fixed in populations, so they do not result in variation at the population level either. Nonetheless, in any population, some amino acid positions will remain constant under purifying selection, others will vary through neutral genetic drift, and still others will differ between species as a result of positive selection for change. How can these evolutionary processes be distinguished?