Flower structure has evolved over time
The flowers of the earliest-diverging clades of angiosperms have a large and variable number of tepals (undifferentiated sepals and petals), carpels, and stamens (Figure 28.11A). Evolutionary change within the angiosperms has included some striking modifications of this early condition: reductions in the number of each type of floral organ to a fixed number, differentiation of petals from sepals, and changes in symmetry from radial (as in a lily or magnolia) to bilateral (as in a sweet pea or orchid), often accompanied by an extensive fusion of parts (Figure 28.11B).

Figure 28.11 Flower Form and Evolution (A) A water lily shows the major features of early flowers: it is radially symmetrical, and the individual tepals, stamens, and carpels are separate, numerous, and attached at their bases. (B) Orchids such as this Venus slipper have a bilaterally symmetrical structure that evolved much later than radial flower symmetry.
According to one hypothesis, the first carpels to evolve were leaves with marginal sporangia, folded but incompletely closed. Early in angiosperm evolution, carpels fused with one another, forming a single, multichambered ovary (Figure 28.12A). In some flowers, the other floral organs are attached at the top of the ovary rather than at the bottom as in Figure 28.4B. The stamens of the most ancient flowers may have been leaflike (Figure 28.12B), with little resemblance to the stamens of the generalized flower seen in Figure 28.4B.
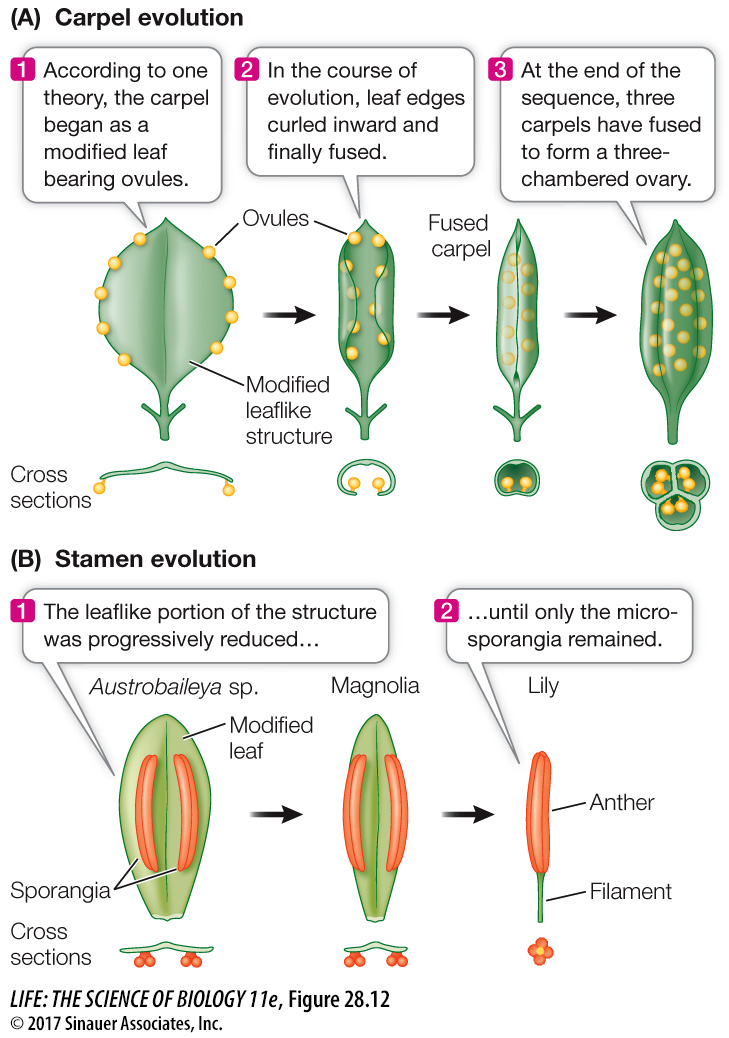
Figure 28.12 Carpels and Stamens Evolved from Leaflike Structures (A) Possible stages in the evolution of a carpel from a more leaflike structure. (B) The stamens of three modern plants show three possible stages in the evolution of that organ. (It is not implied that these species evolved from one another; their structures simply illustrate the possible stages.)
Why do so many flowers have pistils with long styles and anthers with long filaments? Natural selection has favored length in both of these floral organs, probably because length increases the likelihood of successful pollination. Long filaments may bring the anthers into contact with insect bodies, or they may place the anthers in a better position to catch the wind. Similar arguments apply to long styles.
A perfect flower represents a compromise of sorts. On the one hand, by attracting a pollinating bird or insect, the plant is attending to both its female and male functions with a single flower type, whereas plants with imperfect flowers must create that attraction twice—once for each type of flower. On the other hand, the perfect flower can result in self-pollination, which is usually disadvantageous. Another potential problem is that the female and male functions might interfere with each other—for example, the stigma might be placed so as to make it difficult for pollinators to reach the anthers, thus reducing the export of pollen to other flowers.
Might there be a way around these problems? One solution is seen in the bush monkeyflower (Mimulus aurantiacus), which is pollinated by hummingbirds. Its flower has a stigma that initially serves as a screen, hiding the anthers (Figure 28.13). Once a hummingbird touches the stigma, however, one of the stigma’s two lobes is retracted, so that subsequent hummingbird visitors pick up pollen from the previously screened anthers. Thus the first bird to visit the flower transfers pollen from another plant to the stigma. Later visitors pick up pollen from the now-accessible anthers, fulfilling the flower’s male function. Figure 28.14 describes the experiment that revealed the function of this mechanism.
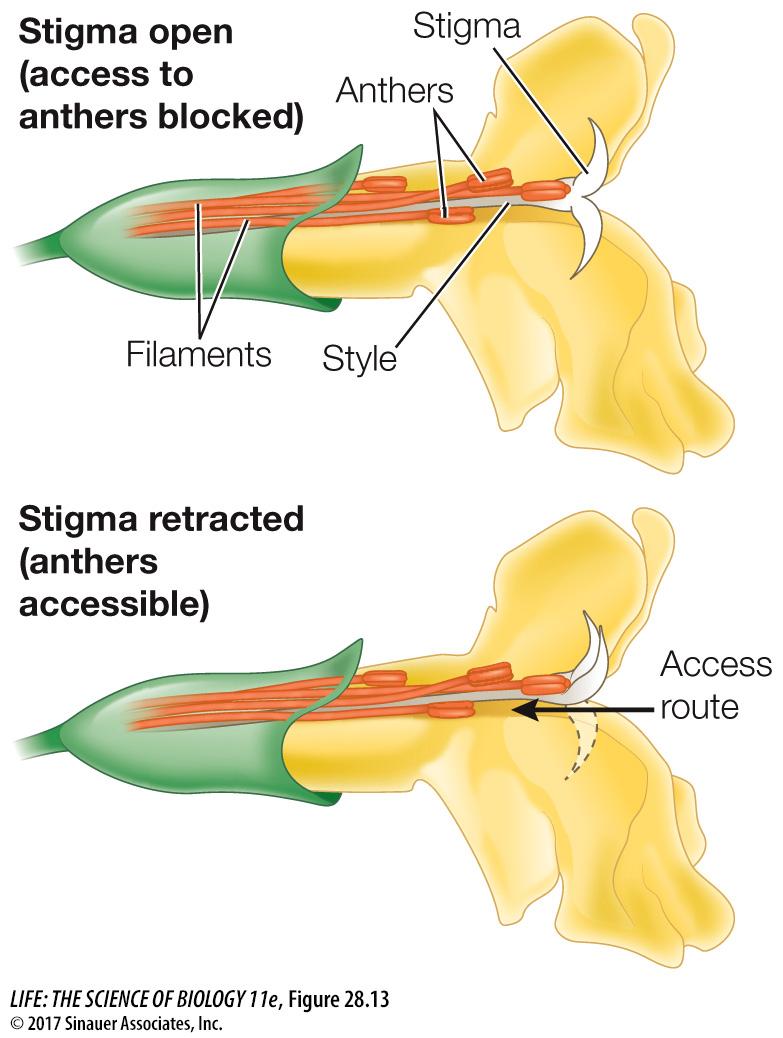
Figure 28.13 An Unusual Way to Prevent Selfing Both long stamens and long styles facilitate cross-pollination, but if these male and female structures are too close to each other, the likelihood of (disadvantageous) self-pollination increases. In Mimulus aurantiacus, the stigma is initially open, blocking access to the anthers. A hummingbird’s touch as it deposits pollen on the stigma causes one lobe of the stigma to retract, creating a path to the anthers and allowing pollen dispersal by subsequent hummingbird visitors.
experiment
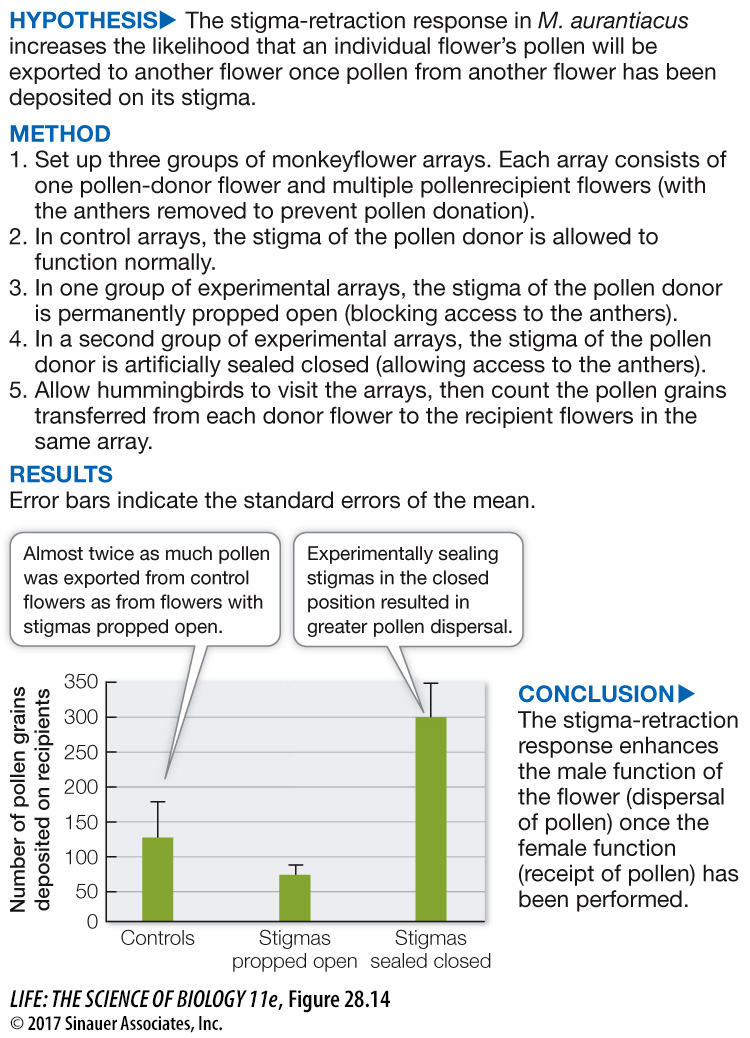
Figure 28.14 The Effect of Stigma Retraction in Monkeyflowers
Original Paper: Fetscher, A. E. 2001. Resolution of male-female conflict in an hermaphroditic flower. Proceedings of the Royal Society B 268: 525–528.
Elizabeth Fetscher’s experiments showed that the unusual stigma retraction response to pollination in monkeyflowers (illustrated in Figure 28.13) enhances the dispersal of pollen to other monkeyflowers.