Gametogenesis produces eggs and sperm
Gametogenesis occurs in the gonads: male testes (singular testis) and female ovaries (singular ovary). The tiny male gametes, sperm, move by beating their flagella. The larger female gametes, eggs or ova (singular ovum), are nonmotile.
Gametes are produced from germ cells, which have their origin in early cell divisions of the embryo and remain distinct from all the other cells of the body (the somatic cells). Germ cells are sequestered in the body of the embryo until its gonads begin to form. The germ cells then migrate to the developing gonads, where they take up residence and proliferate by mitosis, producing spermatogonia (singular spermatogonium) in males and oogonia (singular oogonium) in females (Figure 42.4). Spermatogonia and oogonia are diploid, multiply by mitosis, and are stem cells. They are self-
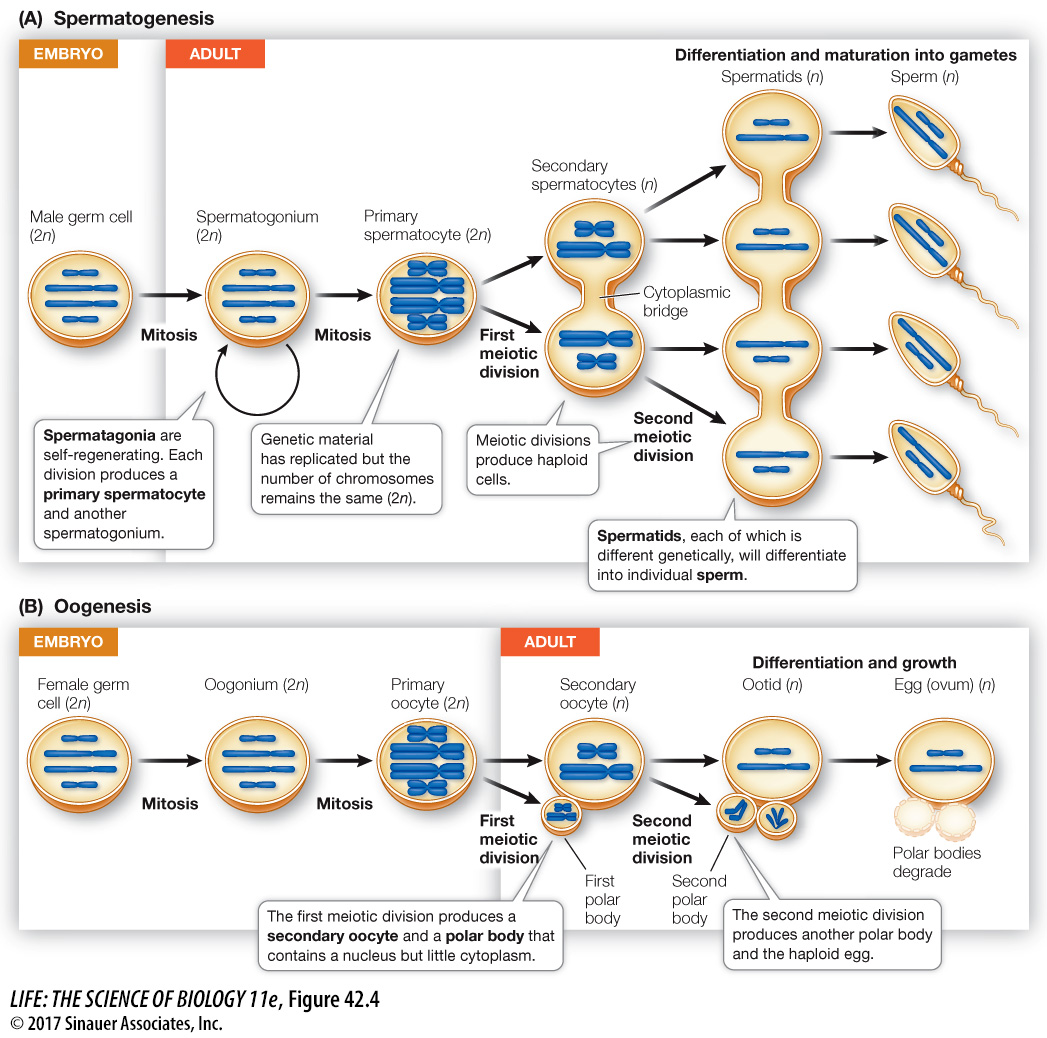
In the next stage of gametogenesis, meiotic cell division reduces the chromosomes to the haploid number (see Key Concept 11.5). The progeny of the spermatogonia and oogonia that enter meiosis are primary spermatocytes and primary oocytes. The steps of meiosis are similar in males and females, but there are important sex differences in gametogenesis.
SPERMATOGENESIS The initial proliferation of male germ cells into spermatogonia proceeds by mitosis in the embryo. But throughout the male life span, spermatogonia continue to divide by mitosis, with one daughter cell retaining the self-
895
One reason that mammalian spermatocytes remain in cytoplasmic contact throughout their development is the asymmetry of sex chromosomes in males. Half the secondary spermatocytes receive an X chromosome, the other half a Y chromosome. The Y chromosome contains fewer genes than the X chromosome, and some of the products of genes found only on the X chromosome are essential for spermatocyte development. By remaining in cytoplasmic contact, all four spermatocytes can share the gene products of the X chromosomes, although only half of them have an X chromosome.
A spermatid bears little resemblance to a mature sperm. Through further differentiation the spermatid becomes compact, streamlined, and developes a flagellum to become motile. We will look at the production of human sperm in Key Concept 42.3.
896
OOGENESIS Oogonia, like spermatogonia, proliferate through mitosis (see Figure 42.4B). The resulting primary oocytes immediately enter prophase of the first meiotic division. In many species, including humans, the oocyte experiences developmental arrest at this point and may remain in that state for days, months, or years. In the human female, this period of arrest is at least 10 years (i.e., until puberty), and some primary oocytes remain in prophase I for over 50 years (i.e., until menopause). In contrast, spermatogenesis continues, uninterrupted, to completion once the primary spermatocyte has differentiated.
During the prolonged prophase I, or shortly before it ends, the primary oocyte grows larger through increased production of ribosomes, RNA, cytoplasmic organelles, and energy stores. At this point the primary oocyte acquires all the energy, raw materials, and RNA that the egg will need to survive its first cell divisions after fertilization. In fact, the nutrients in the egg must maintain the embryo until it is either nourished by the maternal circulatory system or can feed on its own.
When a primary oocyte resumes meiosis, its nucleus completes the first meiotic division near the surface of the cell. The daughter cells of this division receive grossly unequal shares of cytoplasm. This asymmetry represents another major difference from spermatogenesis, in which cytoplasm is apportioned equally. The daughter cell that receives almost all the cytoplasm becomes the secondary oocyte, and the one that receives almost none forms the first polar body (see Figure 42.4B).
The second meiotic division—
A second period of arrested development occurs after the first meiotic division forms the secondary oocyte. The egg may be expelled from the ovary in this condition. In many species, including humans, the second meiotic division is not completed until the egg is fertilized by a sperm.