Fertilization is the union of sperm and egg
The union of the haploid sperm and the haploid egg (fertilization) creates a single diploid cell, a zygote, which will develop into an embryo. Fertilization does more than just restore the full genetic complement of the animal. The processes associated with fertilization help the egg and sperm get together, prevent the union of the sperm and egg of different species, and guarantee that only one sperm will enter and activate the egg. Fertilization involves a complex series of events:
The sperm and the egg chemically recognize each other.
The sperm is activated, enabling it to gain access to the cell membrane of the egg.
The cell membrane of the egg fuses with the cell membrane of a single sperm.
The egg blocks entry of additional sperm.
The egg is metabolically activated and stimulated to start development.
The egg and sperm nuclei fuse to create the diploid nucleus of the zygote.
SPECIFICITY IN SPERM–
Sea urchin eggs release chemical attractants that increase the motility of sperm and cause them to swim toward the egg. These chemical attractants are species-
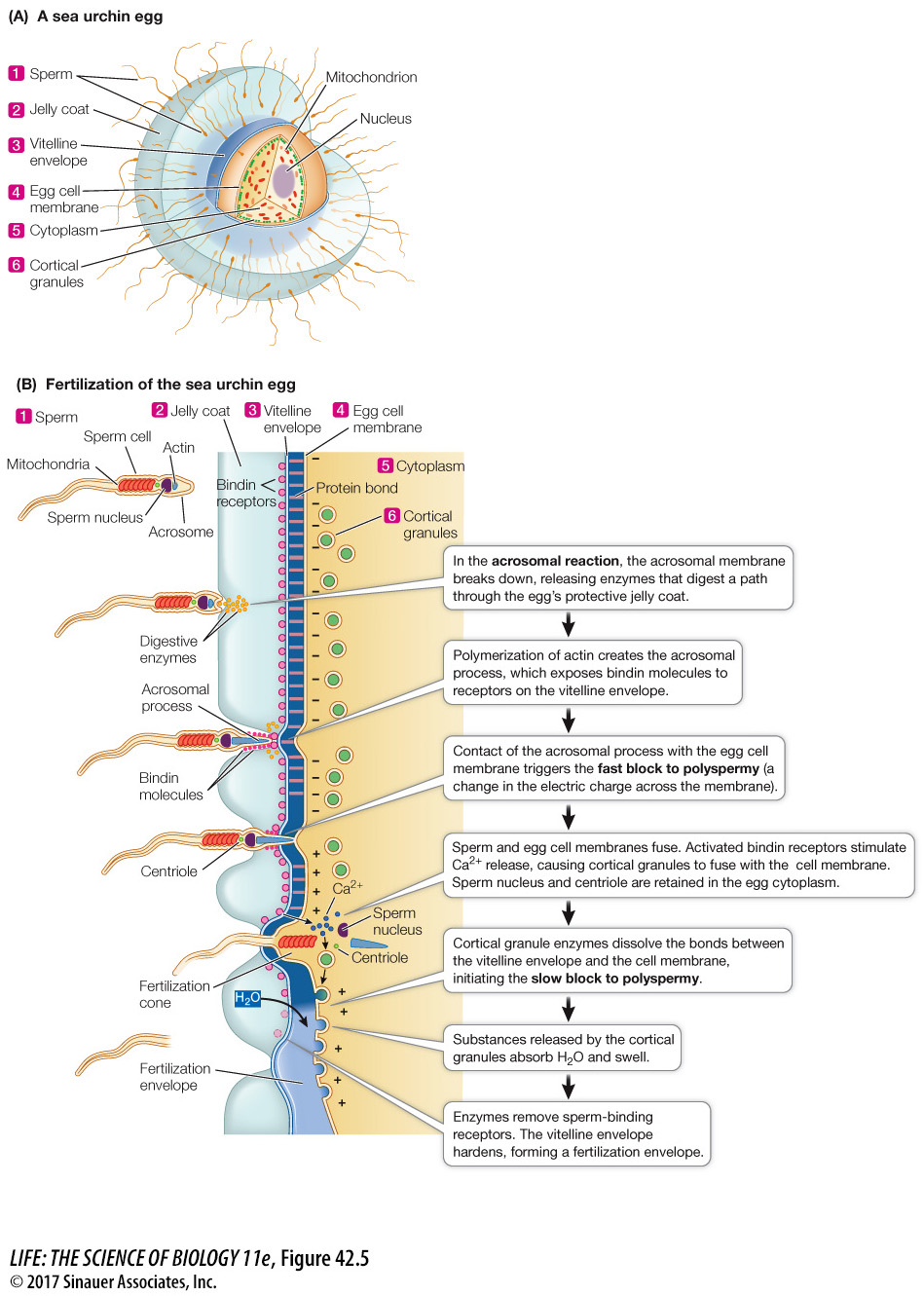
When sperm reach an egg, they must get through two protective layers before they can fuse with the egg cell membrane. The eggs of sea urchins are covered with a jelly coat that surrounds a proteinaceous vitelline envelope (Figure 42.5A). The success of a sperm’s assault on these protective layers depends on a membrane-
Animation 42.1 Fertilization in a Sea Urchin Egg
www.life11e.com/
The acrosome contains enzymes and other proteins. When a sperm makes contact with an egg of its own species, substances in the jelly coat trigger an acrosomal reaction, which begins with the breakdown of the cell membrane covering the sperm head and the underlying acrosomal membrane (Figure 42.5B). The acrosomal enzymes are released and digest a hole through the jelly coat.
As a result of the polymerization of actin triggered by the acrosomal reaction, an acrosomal process extends out of the head of the sperm. The acrosomal process is coated with species-
897
*connect the concepts Because mitochondrial genes are inherited only from the mother, traits controlled by those genes are not inherited in a Mendelian fashion (see Key Concept 12.5), and they enable establishment of matrilineal phylogenies (see Key Concept 21.2).
In animals that practice internal fertilization, mating behaviors help guarantee species specificity, but egg–
Question
Q: What would be the consequences of two sperm fertilizing the egg?
If two sperm fertilized an egg, there would be two sets of male chromosomes in the egg, which would disrupt the subsequent cell divisions.
898
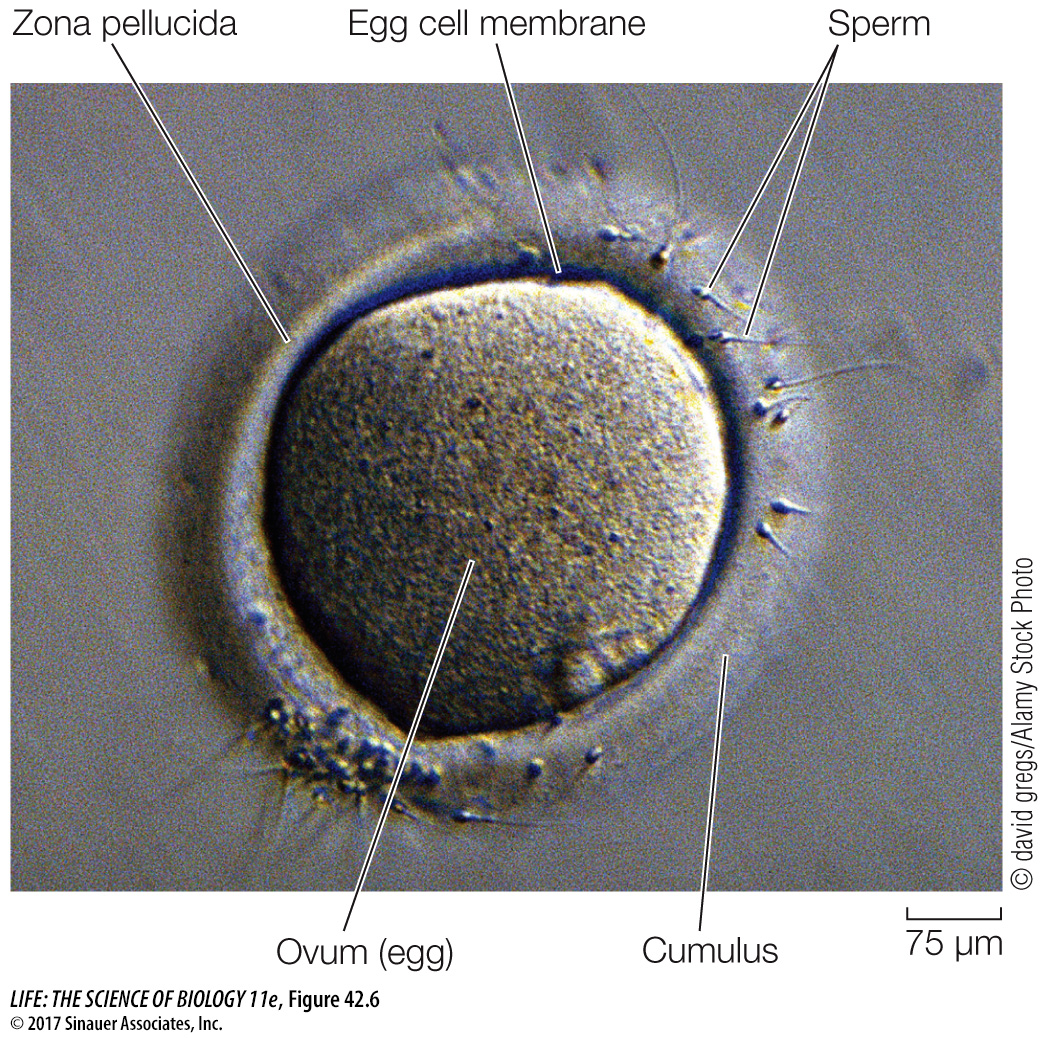
Unlike the jelly coat of sea urchin eggs, the cumulus of mammalian eggs does not trigger the acrosomal reaction. When sperm make contact with the zona pellucida, a species-
BLOCKS TO POLYSPERMY The fusion of the sperm and egg cell membranes and the entry of the sperm into the egg initiate a programmed sequence of events. The first responses to sperm entry are blocks to polyspermy: mechanisms that prevent more than one sperm from entering the egg. Survival of the embryo is unlikely if more than one sperm enters the egg. The sperm contributes a haploid set of chromosomes, so an extra set would disrupt the mitotic division of the egg and subsequent cells. In addition, in most mammals the sperm contributes a centriole to the fertilized egg, and the *centriole forms the centrosome which is critical for organizing the mitotic spindle. Having two centrosomes would disrupt mitosis of the fertilized egg.
*connect the concepts The roles of the centriole and the centrosome in mitosis are discussed in Key Concept 11.3.
Blocks to polyspermy have been studied extensively in sea urchin eggs, which can be fertilized in a dish of seawater. Within seconds after the sperm membrane contacts the egg membrane, an influx of sodium ions changes the electric charge difference across the egg cell membrane. This fast block to polyspermy prevents the fusion of any other sperm with the egg cell membrane, but it is transient. The change in membrane electric charge lasts only about a minute, but that is enough time to allow a slower block to sperm entry to develop.
The slow block to polyspermy involves converting the vitelline envelope to a physical barrier that sperm cannot penetrate. Before fertilization, the vitelline envelope is bonded to the egg cell membrane. Just under the cell membrane are vesicles called cortical granules (see Figure 42.5) which contain enzymes and other proteins.
The sea urchin egg, like all animal cells, sequesters calcium in its endoplasmic reticulum. Sperm entry into the egg stimulates the release of calcium ions from the endoplasmic reticulum and into the egg cytosol. This increase in cytosolic calcium causes the egg’s cortical granules to fuse with the cell membrane and release their contents. Cortical granule enzymes break the bonds between the vitelline envelope and the cell membrane, and other proteins released from the cortical granules attract water into the space between them. As a result, the vitelline envelope rises to form a fertilization envelope. Cortical granule enzymes also degrade sperm-
In mammalian eggs, sperm entry does not cause a rapid change in membrane potential, but it does trigger a release of calcium from the endoplasmic reticulum. As in the sea urchin egg, increased calcium causes the cortical granules to fuse egg with the egg cell membrane. A fertilization envelope does not form around the mammalian egg, but the cortical granule enzymes destroy the sperm-