Invagination at the vegetal pole initiates gastrulation in the sea urchin
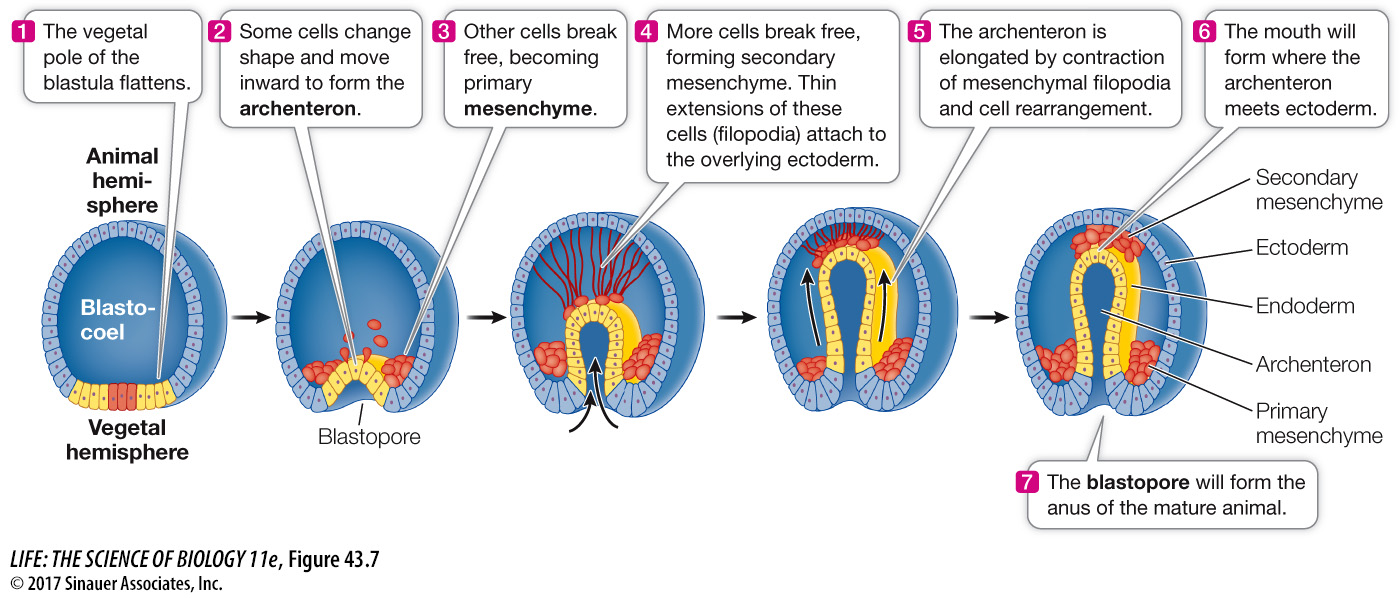
The sea urchin blastula is a hollow ball of cells only one cell layer thick. At the end of blastulation the rate of mitosis slows, and the beginning of gastrulation is marked by a flattening of the vegetal hemisphere (Figure 43.7). Some cells at the vegetal pole break away from neighboring cells and migrate into the cavity to become mesenchyme—cells of the middle germ layer, the mesoderm. Mesenchymal cells are not organized in tightly packed sheets or tubes like epithelial cells are; they act as independent units, migrating into and among the other tissue layers. These early mesenchymal cells will contribute to the skeletal structures of the sea urchin.
Question
Q:How do cells on the surface of the vegetal pole become the cells lining the gut?
Gastrulation involves the involution of vegetal pole cells over the lip of the blastopore to form the archenteron thus forming the lining of the gut.
924
The flattening at the vegetal pole results from changes in the shape of individual blastomeres. These cells, which are originally rather cuboidal, become wedge-
We should recall at this point that where the mouth and anus form during gastrulation distinguishes the two major groups of animals that have bilateral symmetry, the protostomes and the deuterostomes (see Key Concept 30.1). If the blastopore becomes the mouth, the animals are classified as protostomes (“mouth first”), and if the blastopore becomes the anus, the animals are classified as deuterostomes (“mouth second”).
What mechanisms control the various cell movements of sea urchin gastrulation? The immediate answer is that specific properties of particular blastomeres change. For example, some vegetal cells change shape and bulge into the blastocoel, and these cells become mesenchyme. Once they lose contact with their neighboring cells on the surface of the blastula, they send out filopodia that then move along an extracellular matrix of proteins laid down by the cells lining the blastocoel.
A deeper understanding of gastrulation requires that we discover the molecular mechanisms whereby different blastomeres develop different properties. Cleavage systematically divides the cytoplasm of the egg. The sea urchin blastula at the 64-
If different tiers of blastula cells are separated experimentally, they show different developmental potentials; only cells from the vegetal pole are capable of initiating the development of a complete larva. It has been proposed that these differences are due to uneven distribution of various transcriptional regulatory proteins in the egg cytoplasm. As cleavage progresses, these proteins end up in different groups of cells. Therefore specific sets of genes are activated in different cells, determining their different developmental capacities.
Next we will turn to gastrulation in the frog and to the key signaling molecules involved.