Rod and cone cells are the photoreceptors of the vertebrate retina
The photoreceptors of the vertebrate retina are two types of modified neurons called rod cells and cone cells based on their shapes (Figure 45.16). Rod cells are highly light-sensitive and perceive shades of gray in dim light. Cone cells function at high light levels and are responsible for high-acuity color vision. The human retina has about 5 million cones and 100 million rods, but the density of each varies across the retina.
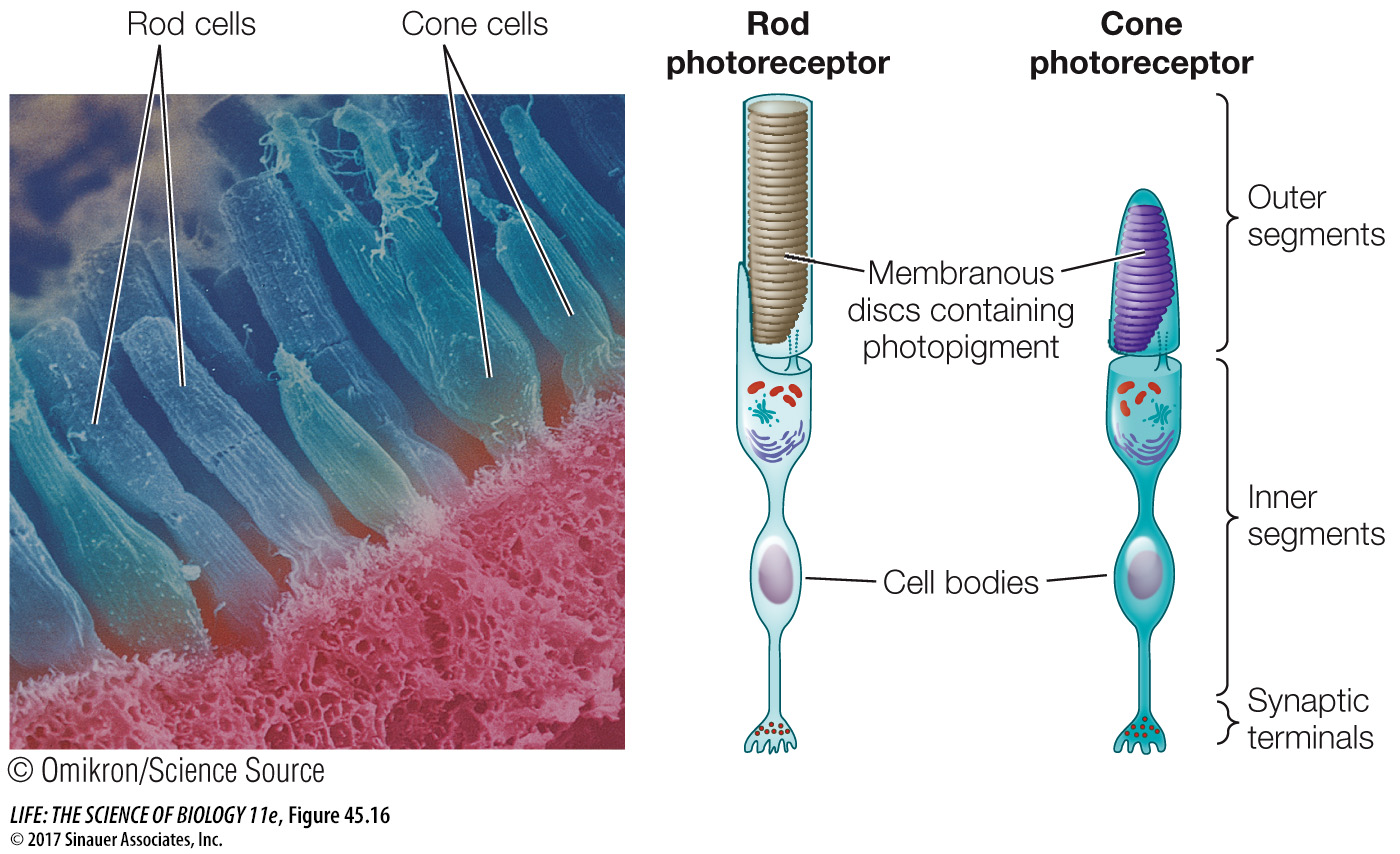
Figure 45.16 Rods and Cones This scanning electron micrograph of photoreceptors in the retina of a mud puppy (an amphibian) shows cylindrical rods and tapered cones.
ROD CELLS Rod cells do not produce action potentials, but they release neurotransmitter from their bases, where they form synapses with the next neurons in the visual pathway. Each rod cell has an outer segment, an inner segment, and a synaptic terminal (see Figure 45.16). The highly specialized outer segment contains stacks of discs made up of cell membranes densely packed with rhodopsin. These discs capture photons of light. The inner segment contains the cell nucleus, mitochondria, and other organelles. The synaptic terminal is where the rod cell communicates with other neurons.
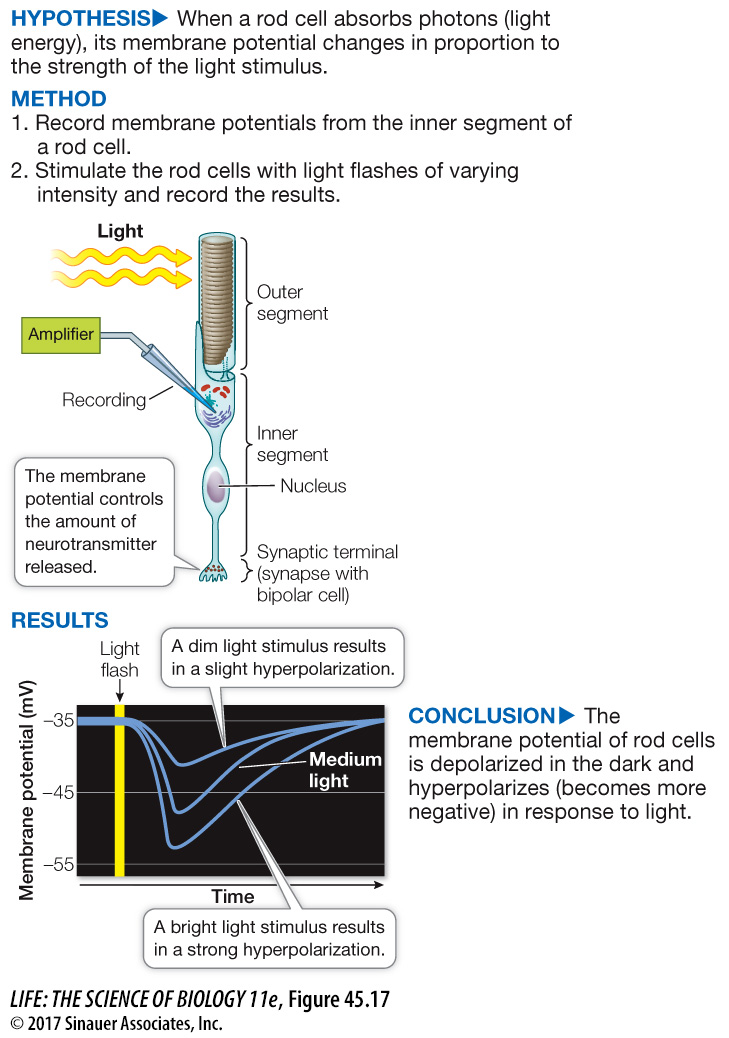
To see how a rod cell responds to light, we can penetrate a single rod cell with an electrode and record its membrane potential in the dark and in the light, as shown in Figure 45.17. From what you have learned about other types of sensory receptors, you might expect that stimulation of the rod cell by light would make its membrane potential less negative, but the opposite is true—it becomes more negative, that is, it becomes hyperpolarized.
experiment
Figure 45.17 How Light Stimulates Rod Cells
Original Paper: Baylor, D. A. and B. J. Nunn. 1986. Electrical properties of the light-sensitive conductance of rods of the salamander Ambystoma tigrinum. Journal of Physiology 371: 115–145.
The cell membrane of a rod cell hyperpolarizes—becomes more negative—in response to a flash of light. Rod cells do not fire action potentials, but in response to the absorption of light energy, the neuron undergoes a change in membrane potential.
Rod cell outer segments have cGMP gated sodium channels. In the dark, abundant cGMP in the cell keeps these channels open, creating the dark current. When light is flashed on the dark-adapted rod cell, these Na+ channels close in proportion to the intensity of the light, and the cell’s membrane potential becomes more negative—it hyperpolarizes (see Figure 45.17). The rate of neurotransmitter release from the base of the cell changes as membrane potential changes. As the rod cell hyperpolarizes, its release of neurotransmitter decreases.
How does the absorption of light by rhodopsin hyperpolarize the rod cell? As illustrated in Figure 45.15, when rhodopsin is excited by light it undergoes conformational changes that initiate a signaling cascade. The dark-adapted rod cell has open Na+ channels—the depolarizing dark current (Figure 45.18). Light photons excite rhodopsin, which activates a G protein called transducin. Activated transducin activates a phosphodiesterase (PDE). Activated PDE converts cyclic GMP (cGMP) to GMP, which causes the cGMP gated Na+ channels to close, changing the balance between the inward flow of Na+ and the outward flow of K+ and causing the cell to hyperpolarize.
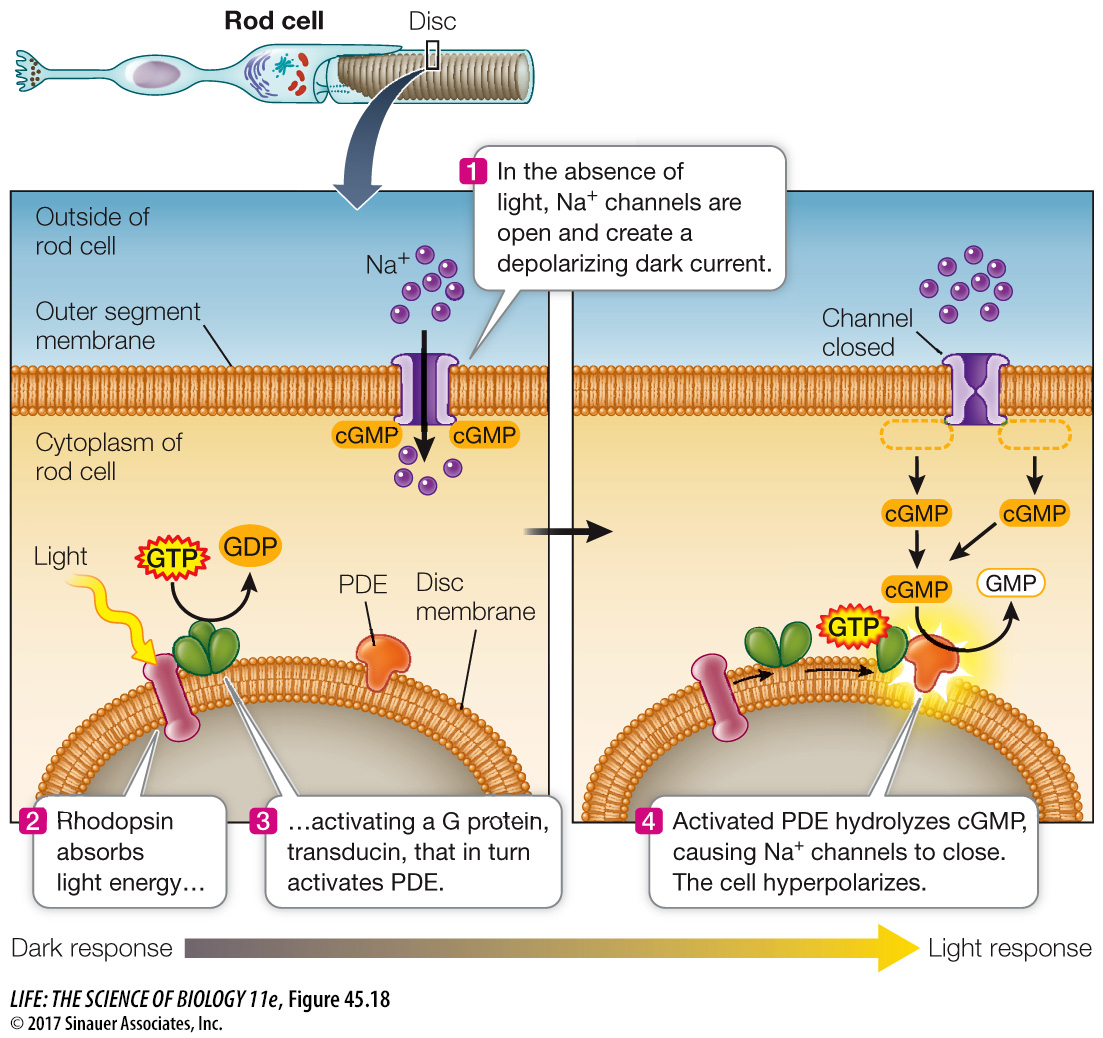
Figure 45.18 Light Absorption Closes Sodium Channels The absorption of light by rhodopsin initiates a signaling cascade that hyperpolarizes the rod cell.
This cascade may seem like a roundabout way of doing business, but its advantage is its enormous amplification ability. Each molecule of light-excited rhodopsin can activate several hundred transducin molecules, thus activating a large number of PDE molecules. The catalytic capacity of PDE is great—one PDE molecule can hydrolyze several hundred molecules of cGMP per second. The bottom line is that a single photon of light can result in the closure of a huge number of Na+ channels.
CONE CELLS Cone cells are responsible for the high-acuity color vision of day-active vertebrates such as humans. It is therefore logical that the highest density of cone cells is in the area of the retina that receives light from the center of the visual field, a region called the fovea (see Figure 45.13A). The human fovea has about 160,000 cones per square millimeter. But humans are not the champions of high-acuity vision; a hawk’s fovea has almost twice that number of cones, making the hawk’s vision much sharper than ours. Birds also have two foveae in each eye; one receives light from straight ahead, the other from a more lateral field of vision. The forward-looking foveae make binocular vision possible, while the lateral-looking foveae provide high-acuity vision. Birds use both sets of foveae by frequently turning their heads slightly; they cannot move their eyes in the sockets as humans can.
Cones have low sensitivity to light and contribute little to night vision. Night vision depends mostly on rod cells, which is why vision in dim light is mostly in shades of gray and acuity is low. You may have trouble seeing a small object at night when you are looking straight at it—that is, when its image is falling on your fovea. If you look a little to the side, so that the image falls on a rod-rich area of your retina, you see the object better. Astronomers looking for faint objects in the sky learned this trick a long time ago. The retinas of nocturnal animals, such as flying squirrels, contain a high percentage of rods. By contrast, some animals that are active only during the day (such as chipmunks) have mostly cones in their retinas.
The human retina has three kinds of cone cells, each containing slightly different opsin molecules that differ in the wavelengths of light they absorb best. Although the same 11-cis-retinal group is the light-absorber in all three kinds of cones, its molecular interactions with opsin determine the spectral sensitivity of the cone cell as a whole (Figure 45.19). Those that absorb maximally around 419 nanometers (nm) are called S-cones for short wavelength (blue range). Those that absorb maximally around 531 nm are called M-cones for medium wavelength (green range), and those that absorb maximally around 559 nm are called L-cones for long wavelength (yellow-red range). Because different wavelengths of light are differentially absorbed by the different cone cell visual pigments, the brain interprets the relative inputs from the different classes as a full range of color. Color blindness in humans results from the absence or dysfunction of one or more of the three classes of cone cells. Some mammals have only one or two classes of cone cells, whereas birds have four.
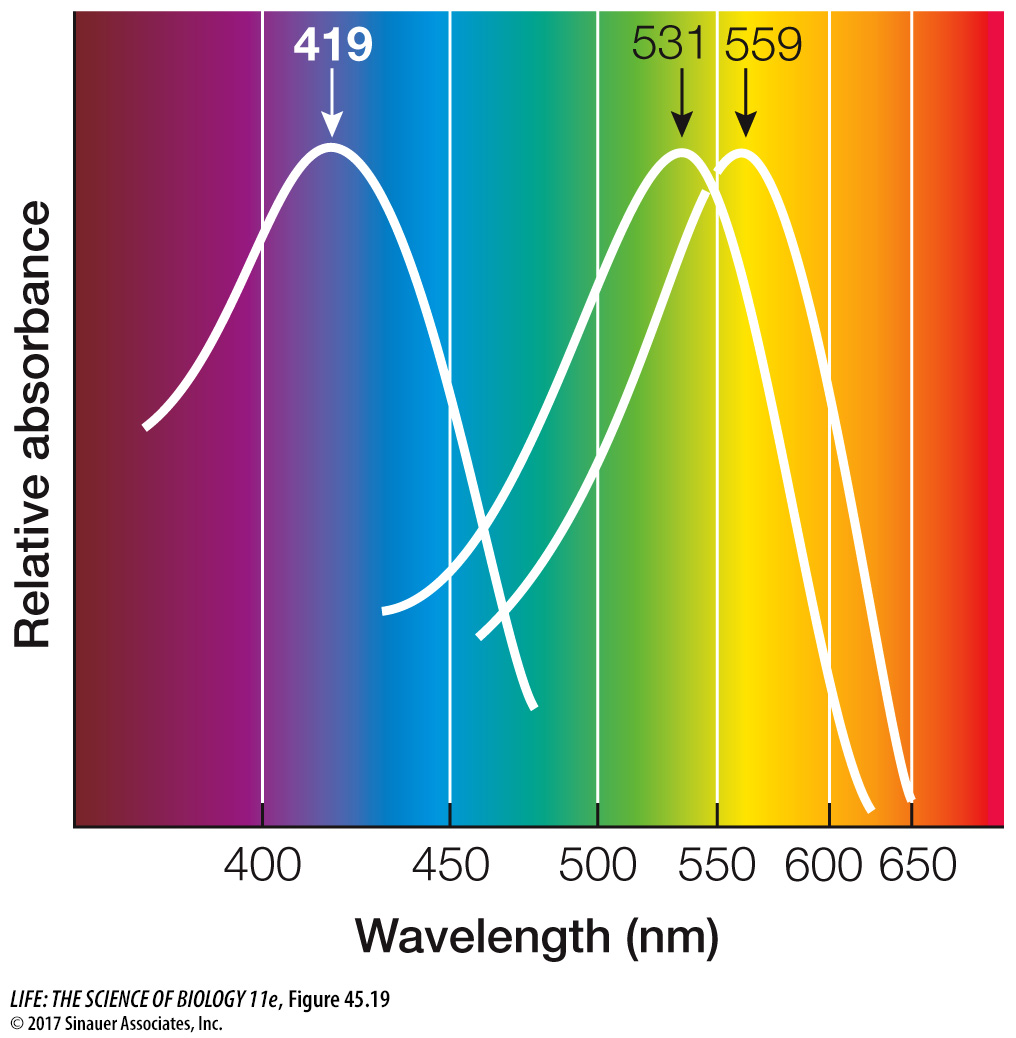
Figure 45.19 Absorption Spectra of Cone Cells The three kinds of cone cells contain slightly different visual pigments that absorb different wavelengths of light.