The cerebrum is responsible for complex behavior and consciousness
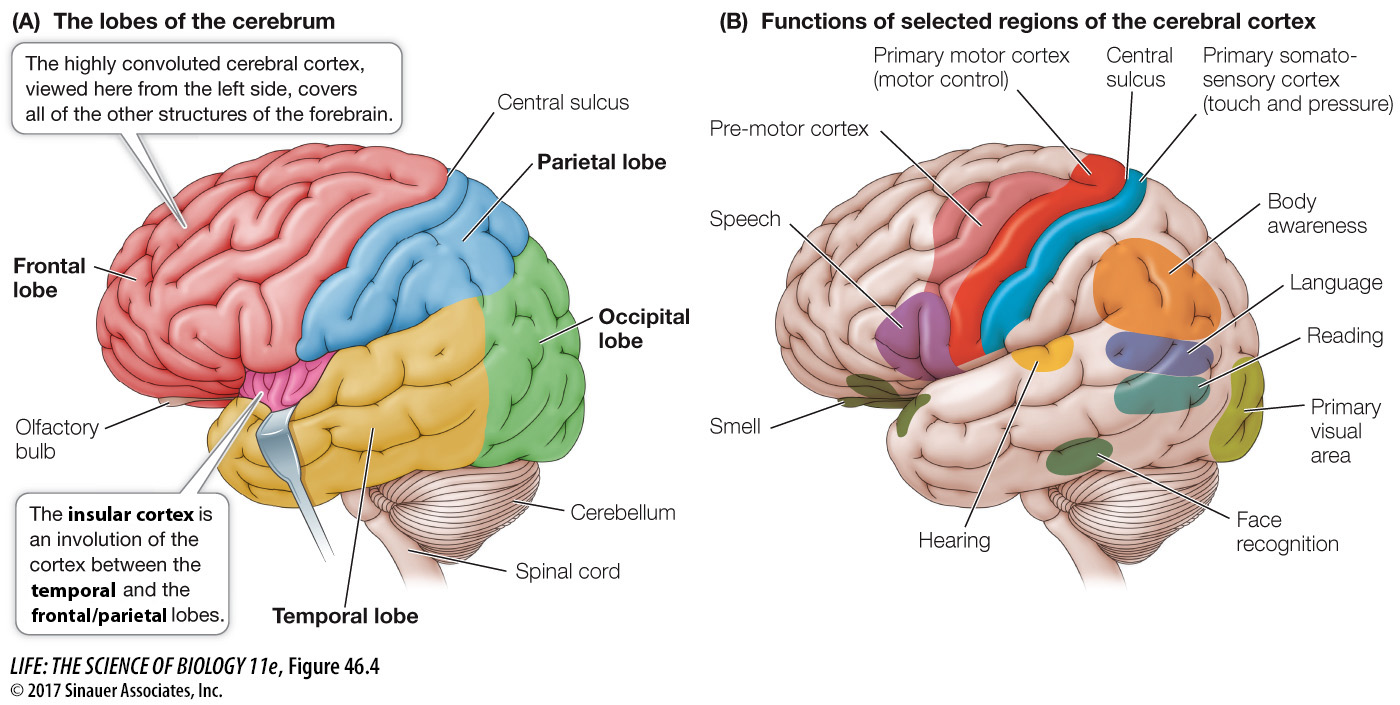
The cerebrum is the dominant structure in the mammalian brain. In humans it is so large that it covers all other parts of the brain except the cerebellum (Figure 46.4A). Layers of cells at the surface of the cerebrum form the cerebral cortex, which is about 4 millimeters thick and is folded into ridges (gyri; singular gyrus) and valleys (sulci; singular sulcus). If the cortex were flattened out, it would be about 1 square meter, but the foldings, or convolutions, enable that extensive surface area of cortex to fit within the skull.
Activity 46.1 The Human Cerebrum
www.life11e.com/
As we explore the infoldings of the cerebral cortex and other parts of the brain, we will occasionally mention an individual whose brain was damaged by an accident or other unfortunate event. Until recently the study of such individuals has been the main source of functional information about the human brain, but new imaging technologies such as positron emission tomography (PET) and magnetic resonance imaging (MRI) are providing a wealth of new information and opportunities to study the human brain.
A curious feature of the human nervous system is that the left side of the body is served (in both sensory and motor aspects) mostly by the right side of the brain, and the right side of the body is served mostly by the left side of the brain. Thus sensory input from the right hand goes to the left cerebral hemisphere, and sensory input from the left hand goes to the right cerebral hemisphere. The exception is the head, where the left side is controlled by the left cerebral hemisphere and the right side by the right cerebral hemisphere. The two hemispheres are not symmetrical with respect to all functions. Language abilities, for example, reside predominantly in the left hemisphere in most people.
Different regions of the cerebral cortex have specific functions (Figure 46.4B). Some of those functions are easily defined, such as receiving and processing sensory information or generating motor commands, but in most humans most of the cortex is involved in higher-
To understand the cerebral cortex, it helps to have an anatomical road map. Viewed from the left side, the left cerebral hemisphere looks like a boxing glove for the right hand with the fingers pointing forward, the thumb pointing out, and the wrist at the rear. The “thumb” area is the temporal lobe, the fingers the frontal lobe, the back of the hand the parietal lobe, and the wrist the occipital lobe (see Figure 46.4A). What you can’t see from the external view is the insular cortex, which is folded in between the thumb and finger areas of the boxing glove (between the frontal/parietal and the temporal lobes). The right cerebral hemisphere shows a mirror image of this arrangement. We will look at each lobe separately.
985
investigating life
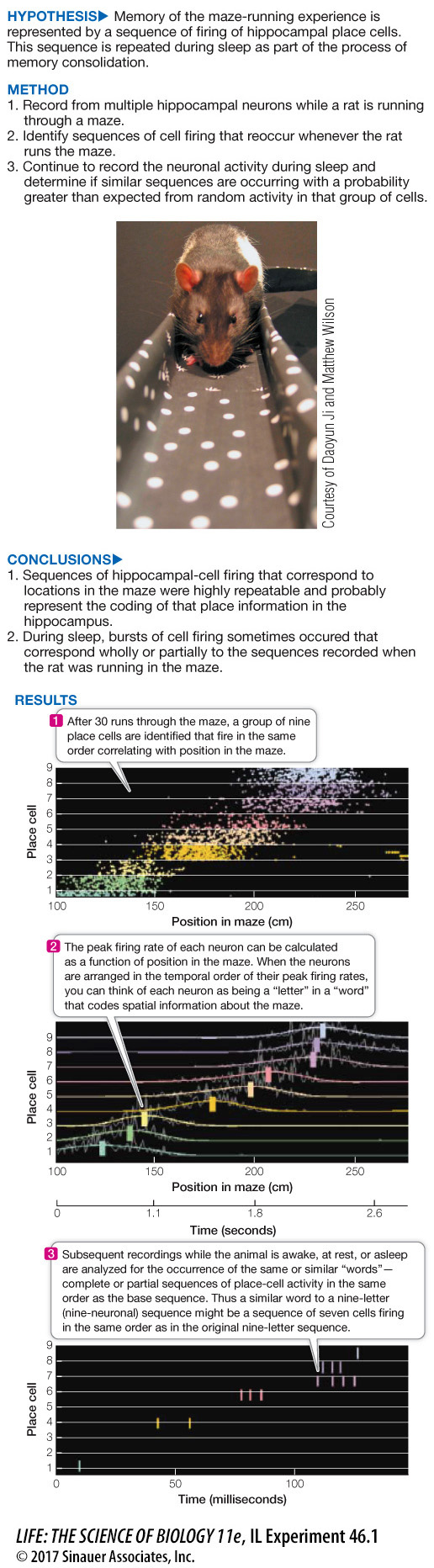
Place Cells Reveal Processes of Memory Consolidation during Sleep
experiment
Original Paper: Lee, A. K. and M. A. Wilson. 2002. Memory of sequential experience in the hippocampus during slow wave sleep. Neuron 36: 1183–
Multiple hippocampal cells were recorded while a rat ran a maze. Individual cells fired only when the rat was at particular locations in the maze; thus the rat’s progress through the maze can be represented by a specific sequence of firing of these “place cells.” Since memory is consolidated during sleep, are the sequences of place cells expressed during the maze-
work with the data
986
To conclude that a particular sequence of cell firing observed during sleep is a significant event—
What is the minimum number of elements that would have to match the baseline sequence for it to be considered a significant event with P < 0.01? To find out, we need to calculate the probabilities of the recorded neurons firing randomly and creating the sequence matches. First, let’s consider the probability of a recurrence by chance of the exact nine-
9! = 9 × 8 × 7 × 6 × 5 × 4 × 3 × 2 × 1 = 362,880
In other words, the probability of repeating the entire nine-
QUESTIONS
Question 1
What is the probability of the sequence 1, 4, 6, 7, 8, 9 out of a random firing of nine cells?
The probability of this sequence out of a random firing of nine cells is first a function of the probability of that one combination from all of the combinations of cells that could constitute six elements of the nine possible. That would be 1/(9 × 8 × 7 × 6 × 5 × 4) = 1/60,480 = 1.6 × 10–5. Then, for the sequence, what is the probability that it would be in the correct order? There is one possible correct order of those six elements in 6 × 5 × 4 × 3 × 2 × 1 possible combinations or 1 in 720, thus the probability would be 1.4 × 10–3. So the probability of recording those specific six elements out of nine in the correct order would be 2.24 × 10–8.
Question 2
What is the probability of a sequence of four letters occurring out of a random firing of nine cells?
The probability of recording four elements in the sequence in the proper order would be: 1/(9 × 8 × 7 × 6) = 1/4,698 = 2.1 × 10–4, and the probability of getting those four elements in the proper order would be 1/24. Thus the probability of recording those specific four elements out of nine in the correct order would be 2.1 × 10–4 × 0.042 = 0.88 × 10–3.
Question 3
Are both of the recordings from Questions 1 and 2 low-
Both of these recordings have a probability of < 0.01, so they would be considered significant low-
Question 4
What do you note about the time frame of the sequence during sleep versus during wakefulness?
The time frame of the sequence recording during sleep is about five times faster than the occurrence of that sequence during wakefulness when the rat is running the maze.
A similar work with the data exercise may be assigned in LaunchPad.
987
THE TEMPORAL LOBE The upper region of the temporal lobe receives and processes auditory information. The association areas of this lobe are involved in recognizing, identifying, and naming objects. Damage to the temporal lobe results in disorders called agnosias, in which the individual is aware of an object but cannot identify it.
Damage to a specific region of the temporal lobe results in the inability to recognize faces. Even old acquaintances cannot be identified by facial features, although they may be identified by other attributes such as voice, body features, and posture. Damage to other association areas of the temporal lobe can cause deficits in understanding spoken language, although speaking, reading, and writing abilities may be intact.
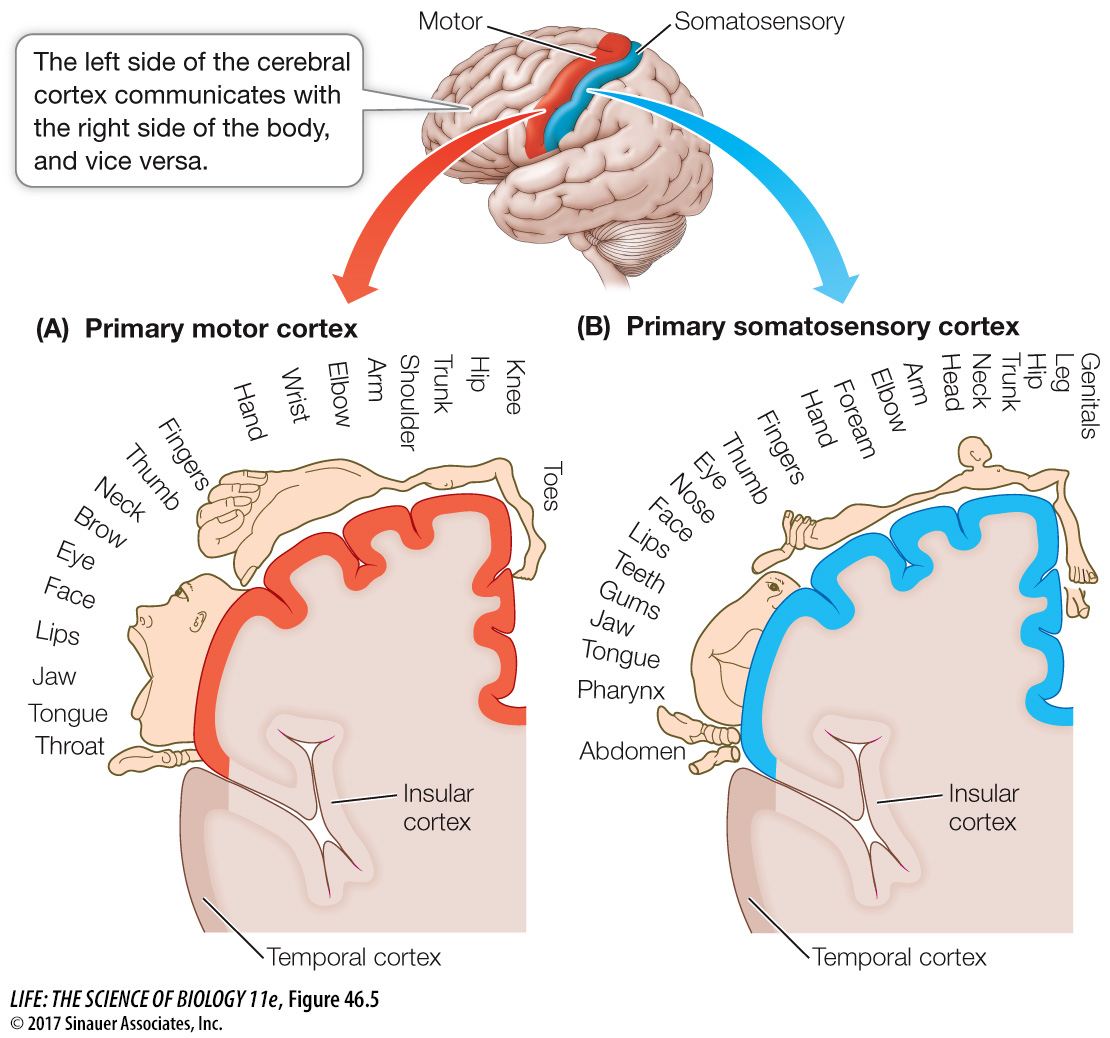
THE FRONTAL LOBE The frontal and parietal lobes are separated by a deep valley called the central sulcus. A strip of the frontal lobe cortex just in front of the central sulcus is called the primary motor cortex (see Figure 46.4B). The neurons in this region control muscles in specific parts of the body; the parts of the body map onto the primary motor cortex, with the head represented in the lower outside region and the legs and feet in the top region near the midline. The graphic drawing of body parts overlying the diagram of the cortex is called a homunculus, and its distortion represents disproportionate innervation. Parts of the body with fine motor control, such as the face and hands, have disproportionate representation (Figure 46.5A). Electrical stimulation of neurons in different regions of the primary motor cortex causes specific muscles of the body to twitch. Just anterior to the primary motor cortex is the pre-
The association functions of the frontal lobe are diverse and best described as having to do with feeling and planning. They are said to have executive function and they contribute significantly to personality. People with frontal lobe damage have drastic alterations of personality and difficulty planning future events. A dramatic case of frontal lobe damage is that of Phineas Gage, who in 1848 was an industrious and responsible young railroad construction foreman. Then a blasting accident shot a meter-
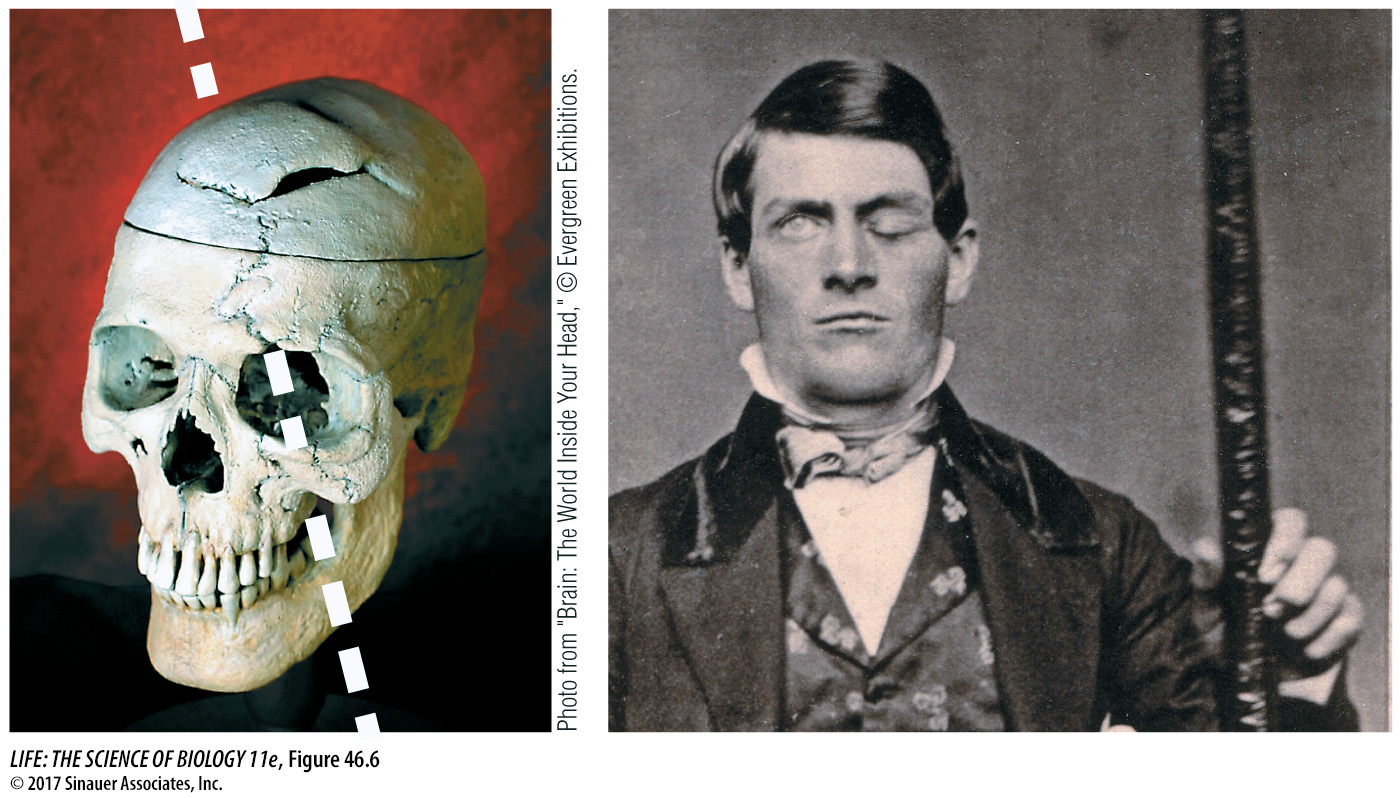
Remarkably, Gage survived, but he had a different personality. In the years following his recovery, he was quarrelsome, impatient, obstinate, and used profane language, which he did not do before. He lost his railroad job and earned money by telling his story and exhibiting his scars (and the tamping iron). Several years later, however, he took a job as a stage coach driver in Chile—
THE PARIETAL LOBE The strip of parietal lobe cortex just behind the central sulcus is the primary somatosensory cortex (see Figure 46.4B). This area receives touch and pressure information relayed from the body through the thalamus.
988
As with the primary motor cortex, the entire body surface can be mapped onto the primary somatosensory cortex (Figure 46.5B). Areas of the body that have a high density of tactile mechanoreceptors and are capable of making fine discriminations in touch (such as the lips and fingers) have disproportionately large representation. If a very small area of the primary somatosensory cortex is stimulated electrically, the subject reports feeling specific sensations, such as touch, in a localized part of the body.
A major association function of the parietal lobe is attending to complex stimuli. Damage to the right parietal lobe causes a condition called contralateral neglect syndrome, in which the individual tends to ignore stimuli from the left side of the body or the left visual field. Such individuals have difficulty performing complex tasks, such as dressing the left side of the body; an afflicted man may not be able to shave the left side of his face. When asked to copy simple drawings, a person who exhibits this syndrome can do well with the right side of the drawing but not the left.
The parietal cortex is not symmetrical with respect to its role in attention. Damage to the left parietal cortex does not cause the same degree of neglect of the right side of the body. You will see similar asymmetries in cortical function later in the chapter when we discuss language.
THE OCCIPITAL LOBE The occipital lobe receives and processes visual information. The association areas of the occipital cortex are essential for making sense of the visual world and translating visual experience into language. Some deficits resulting from damage to these areas are specific. In one case, a woman with limited damage was unable to see motion. Her vision was intact, but she could see a waterfall only as a still image, and an approaching car only as a series of a stationary object at different distances.
THE INSULAR CORTEX The insular cortex, buried deep in the forebrain, receives a great variety of afferent information. The posterior regions receive somatosensory information (e.g., touch, pain, temperature); the middle region combines that with autonomic regulatory information and drives (e.g., hunger, thirst, sex); and the anterior region, which is well developed in higher primates, elephants, and marine mammals, receives information about social interactions. Thus in mammals the insular cortex appears to integrate physiological information from all over the body to create a sensation of how the body “feels,” and in the higher mammals it may extend that function into a sense of self.