Fish gills use countercurrent flow to maximize gas exchange
1027
The internal gills of fish are supported by gill arches that lie between the mouth cavity and the protective opercular flaps on the sides of the fish just behind the eyes (Figure 48.5A). Water flows unidirectionally into the fish’s mouth, over the gills, and out from under the opercular flaps. Thus there is a constant one-
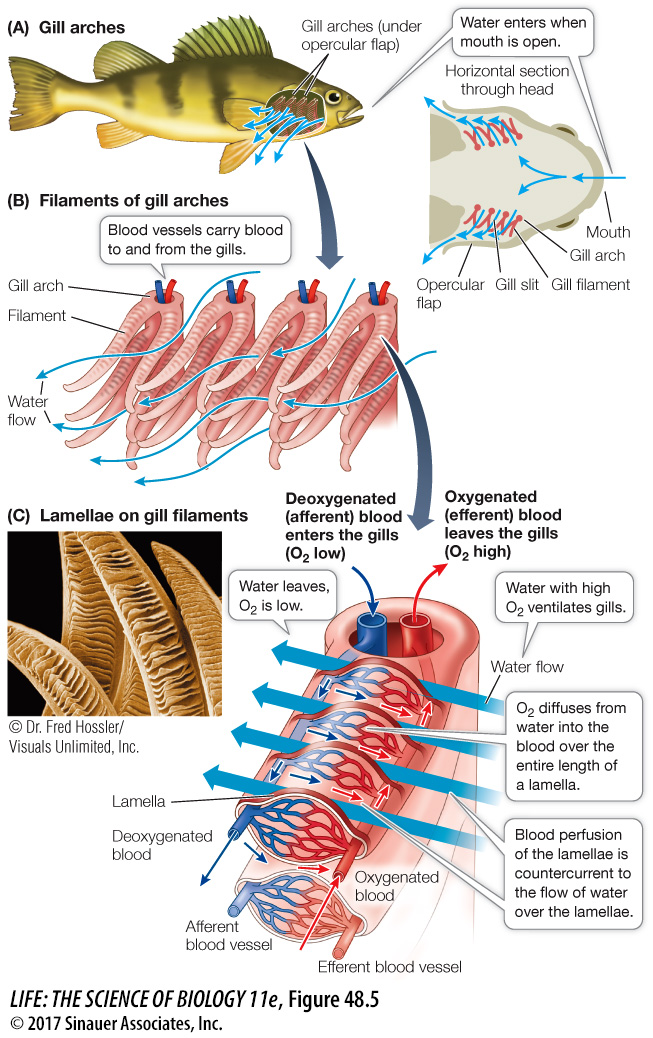
Gills have an enormous surface area for gas exchange because they are so highly subdivided. Each gill consists of hundreds of ribbonlike gill filaments (Figure 48.5B). The upper and lower flat surfaces of each gill filament are covered with rows of evenly spaced folds, or lamellae (Figure 48.5C). The lamellae are the actual gas exchange surfaces. Because the lamellae are exceedingly thin, the path length (L) for diffusion of gases between blood and water is minimized. The surfaces of the lamellae consist of highly flattened epithelial cells, so the water and the fish’s red blood cells are separated by little more than 1–
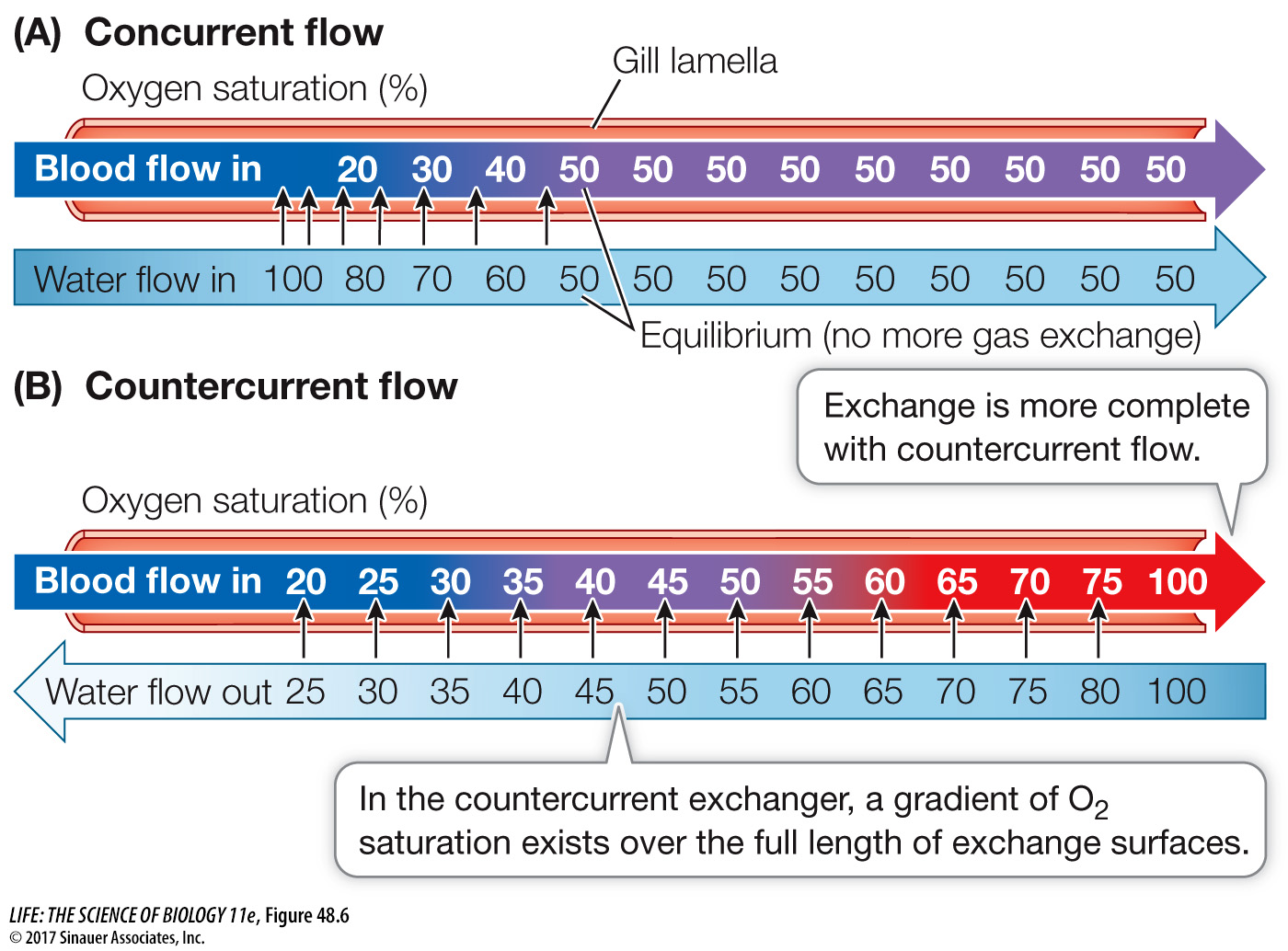
The flow of blood perfusing the inner surfaces of the lamellae, like the flow of water over the gills, is unidirectional. Afferent blood vessels bring deoxygenated blood to the gills, while efferent blood vessels take oxygenated blood away from the gills (see Figure 48.5C). Blood flows through the lamellae in the direction opposite to the flow of water over the lamellae. This countercurrent flow optimizes the PO2 gradient between water and blood, making gas exchange more efficient than it would be in a system using concurrent (parallel) flow (Figure 48.6).
Question
Q: What could make the concurrent exchanger achieve the same maximum O2 saturation as the countercurrent exchanger?
If the flow rate of the water were increased, or the flow rate of the blood were decreased, it would be possible to maximally saturate the blood with a concurrent exchanger. This exchange would be less efficient than countercurrent exchange, however.
Some fishes, including anchovies, tuna, and certain sharks, ventilate their gills by swimming almost constantly with their mouths open. Most fishes, however, ventilate their gills by means of a two-
These adaptations for maximizing the surface area (A) for diffusion, minimizing the path length (L) for diffusion, and maximizing the PO2 gradient allow fishes to extract an adequate supply of O2 from meager environmental sources.