Nephrons have a regular arrangement in the kidney
The kidney is shaped like a kidney bean. When sliced along its long axis on the midline, its key anatomical features are revealed (Figure 51.9B). The ureter and the renal artery and renal vein enter the kidney on its concave (punched-
1103
The organization of nephrons within the kidney is very regular. All of the glomeruli with their Bowman’s capsules are located in the cortex. The initial segments of the renal tubules that come from the Bowman’s capsules are called the proximal convoluted tubules—“proximal” because they are closest to the glomerulus, and “convoluted” because they are twisted (Figure 51.9C). All of the proximal convoluted tubules are located in the cortex.
At the point at which the proximal convoluted tubule enters the medulla, it becomes thin, straight, and descends directly down toward the tip of a renal pyramid. In the medulla the tubule makes a hairpin turn and ascends back to the cortex, forming what is called a loop of Henle. Some nephrons have longer loops of Henle than others. Some 20–
The ascending limb of the loop of Henle becomes the distal convoluted tubule when it reaches the cortex—
The organization of the blood vessels of the kidney closely parallels the organization of the nephrons (see Figure 51.9C). Smaller arteries branch from the renal artery and radiate into the cortex, forming the afferent arterioles that carry blood to each glomerulus. Each glomerulus is drained by an efferent arteriole that gives rise to the peritubular capillaries, most of which surround the proximal and distal convoluted tubules. The intimate associations of the glomerular and peritubular capillaries with the renal tubules enable exchanges between the blood and the specialized regions of the tubules.
Some of the peritubular capillaries run into the medulla in parallel with the loops of Henle and the collecting ducts, forming a vascular network called the vasa recta. All of the peritubular capillaries from a nephron join back together into a venule that joins with venules from other nephrons and eventually leads to the renal vein. As you will see, the concentrating ability of the mammalian kidney depends on water reabsorption in the renal medulla, and the vasa recta are the avenue by which that water gets out of the renal medulla and back into the circulation.
1104
investigating life
How Can Vampire Bats use Blood as Fast Food?
experiment
Original Paper: Busch, C. 1988. Consumption of blood, renal function and utilization of free water by the vampire bat, Desmodus rotundus. Comparative Biochemistry and Physiology Part A: Physiology 90: 141–
You learned at the beginning of the chapter that vampire bats feed once a day. After sunset they fly out, search for a victim, and then return to their roost. This nightly excursion generally takes less than 2 hours. Therefore in a very short time these small animals must take in a large enough volume of their liquid blood diet to supply their energy and water needs for a 24-
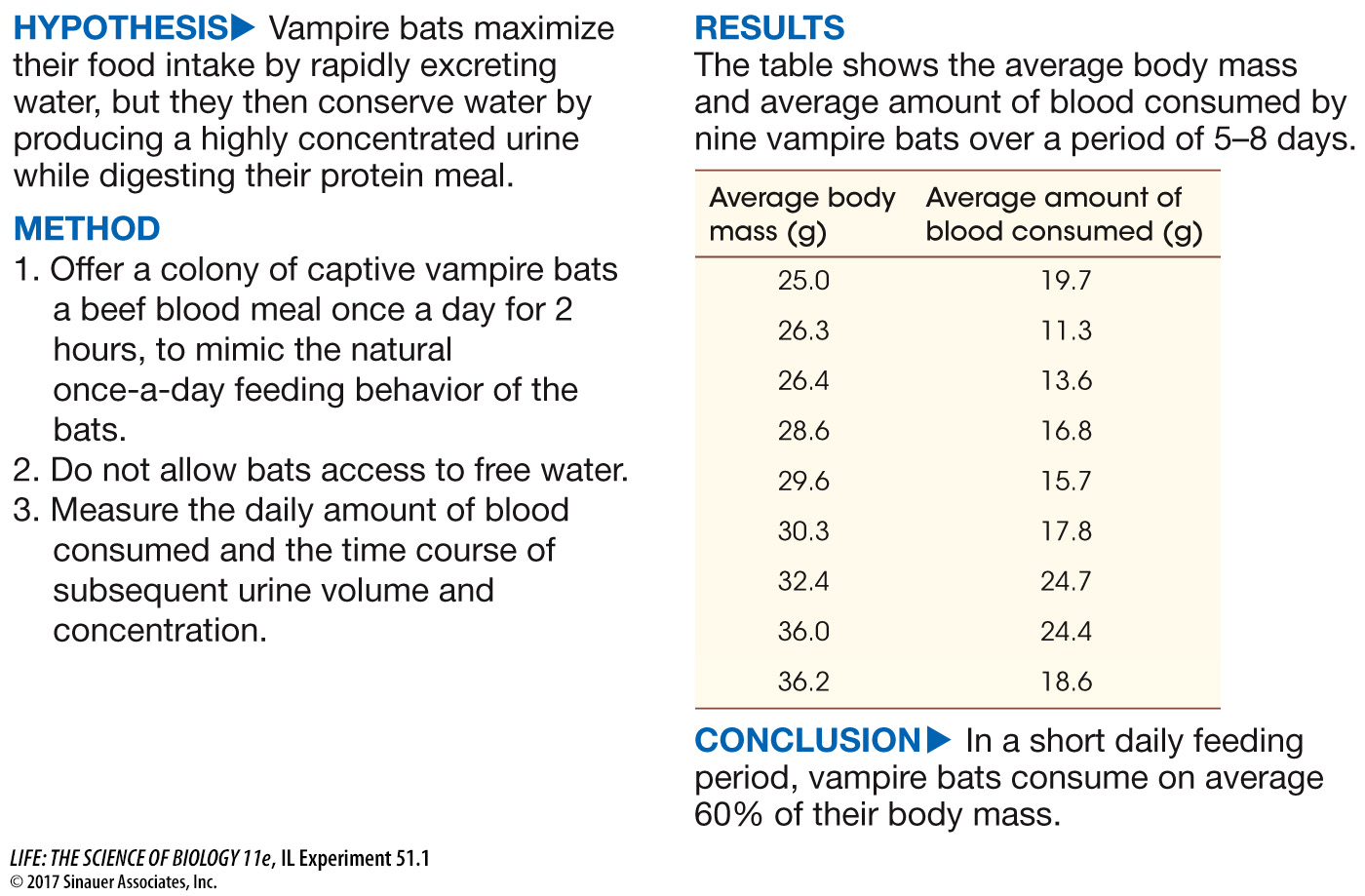
work with the data
To consume 60% of body mass and still be able to fly, the bats must rapidly excrete a large percentage of the water content of their food. The time course of urine flow and its concentration was measured during and after daily feeding for three vampire bats for 1 week and averaged. Create a double plot of the flow and concentration data as a function of time. (—, no data reported.)
QUESTIONS
Question 1
If you averaged the urine flow rates over the first hour following the beginning of the meal, and you assumed that the total blood intake was 60% of body mass and body mass was 30 g, what percentage of the ingested volume would be excreted in that first hour?
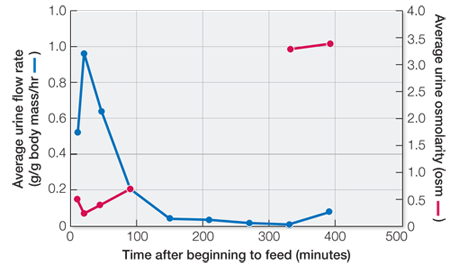
The average urine flow rate over the first hour is 0.18 g/g body mass/hr and for a 30-
Question 2
Considering that the normal osmolarity of interstitial fluid in a mammal is 300 mosm, what is the maximum concentrating factor in these vampire bats?
The maximum concentrating factor in this experiment is 3,400 mosm/300 mosm = 11.3.
Question 3
Three data points are missing in the urine osmolarity curve. Would it be reasonable to plot these data by connecting the 90-
Extrapolating the missing data by connecting the two curves with a straight line would not be reasonable because the urine flow rate falls close to its lowest level at 2.5 hours (150 minutes). It is reasonable to expect that the urine osmolarity would be close to maximum at this time.
| Time after beginning to feed (min) |
Average urine flow rate (g/g body mass/hr) |
Average urine osmolarity (osm) |
|---|---|---|
| 10 | 0.13 | 0.5 |
| 20 | 0.24 | 0.25 |
| 45 | 0.16 | 0.40 |
| 90 | 0.05 | 0.70 |
| 150 | 0.01 | — |
| 210 | 0.01 | — |
| 270 | 0.005 | — |
| 330 | 0.003 | 3.3 |
| 390 | 0.02 | 3.4 |
A similar work with the data exercise may be assigned in LaunchPad.