Biological rhythms coordinate behavior with environmental cycles
Earth turns on its axis once every 24 hours, generating daily cycles of light and dark, temperature, humidity, and tides. In addition, Earth is tilted on its axis, so the light–dark cycle changes as Earth revolves around the sun. These daily and seasonal cycles profoundly influence the physiology and behavior of animals. Animals tend to be active either during the day (diurnal) or at night (nocturnal) and have sensory capabilities appropriate to this distinction. Therefore it is adaptive to organize behavior on a cycle that corresponds with the environmental cycle of light and dark. Similarly, a behavior that is adaptive at one time of year (such as midsummer) may not be adaptive at another time (midwinter). Thus it is important for animals to organize their behavior with respect to time of the day or year and to be able to anticipate those times.
CIRCADIAN RHYTHMS Experimental animals kept under constant conditions with no daily time cues such as light and dark, temperature cycles, or episodic availability of food and water still have daily cycles of activities such as locomotor activity, sleeping, eating, drinking, learning, and just about anything else that can be measured. The persistence of these daily cycles in the absence of environmental time cues suggests that animals have an internal clock. As we mentioned above, because these daily cycles are not exactly 24 hours long, they are known as circadian rhythms.
Biological rhythm can be viewed as a series of cycles, and the length of one of those cycles is the period of the rhythm. Any point in the cycle is a phase of that cycle. When two rhythms completely match, they are in phase, and if a rhythm is shifted (as in the resetting of a clock), it is phase-advanced or phase-delayed. Because the period of a circadian rhythm is not exactly 24 hours, it must be phase-advanced or phase-delayed each day to remain in phase with the daily cycle of the environment. In other words, the rhythm has to be entrained to the environmental cycle of light and dark.
The circadian rhythm of an animal held under constant conditions will run according to its natural period—it will be free-running. If the period is less than 24 hours, the animal will begin its activity a little earlier each day (Focus: Key Figure 52.11). The period of the free-running circadian rhythm is under genetic control. Different species may have different average periods, and within a species, mutations can lead to different period lengths.
focus: key figure
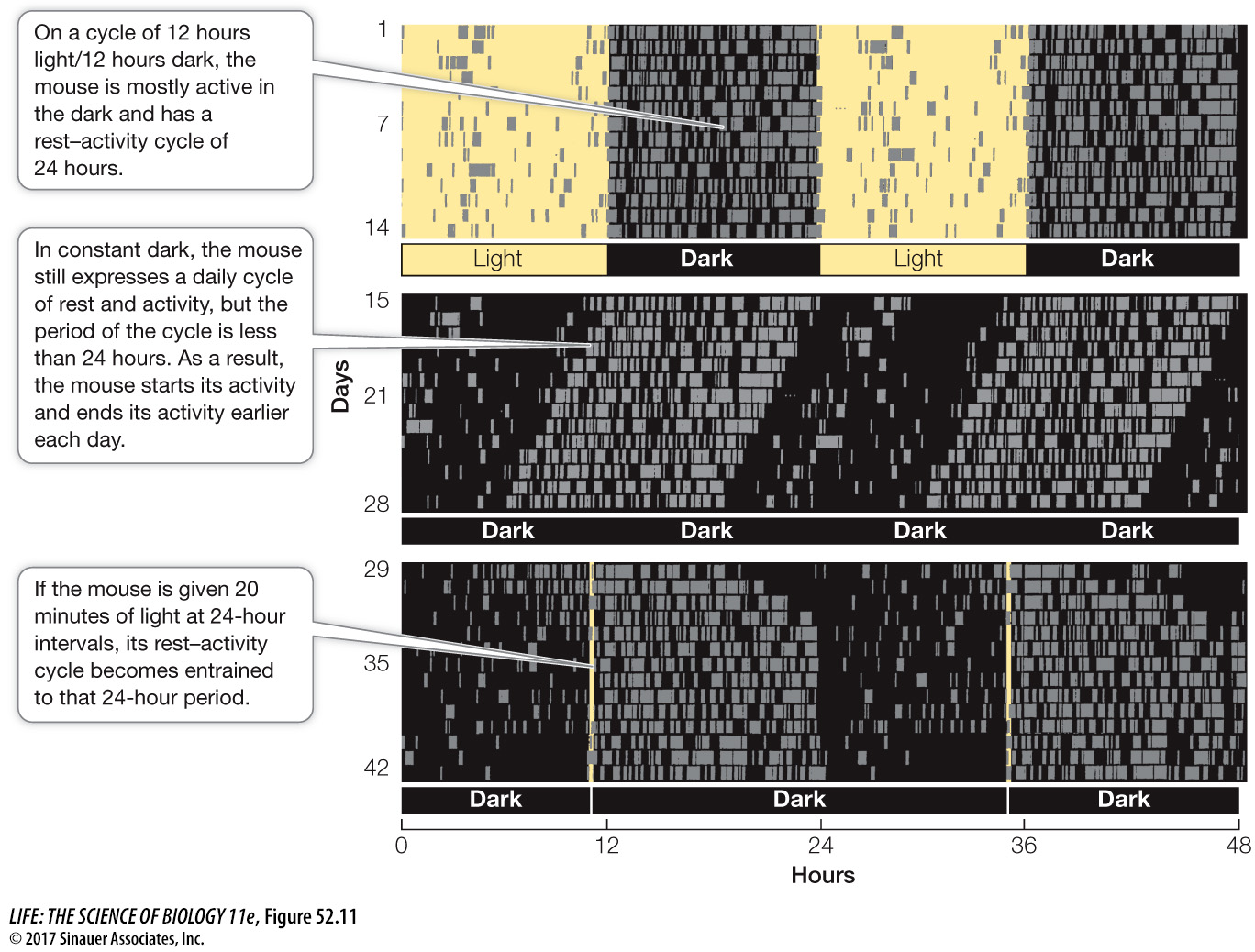
Figure 52.11 Circadian Rhythms Are Entrained by Environmental Cues The activity–rest cycle of a laboratory mouse (a nocturnal animal) responds to the light–dark cycle under which it is kept. The gray bars indicate times when the mouse is running on an activity wheel. Two days of activity are represented on each horizontal line. Thus, day 1 is 0–24 and day 2 is 24–48. Then on the next line day 2 is replotted as 0–24 and day 3 is 24–48, and so on. This double plotting is merely to make the pattern easier to see.
Question
Q: In going from the last activity tracing in the second panel to the light-pulse experiment in the third panel, do this transition and its effects on the circadian clock seem more similar to what you experience if you are recovering from jet lag after traveling from San Francisco to New York, or in the opposite direction?
The extended activity bouts of the mouse in the second panel represent the portion of the animal’s circadian rhythm that corresponds to what its circadian clock is predicting as night. Therefore the light pulse in this experiment would be similar to seeing light early in the night. This is similar to what our experience would be after traveling from east to west—sunset would come later than our circadian clock would predict. In other words, we would get a light stimulus in the early portion of our circadian night. The result would be a delay of the circadian rhythm to bring it in line with the local light-dark cycle.
Animation 52.3 Circadian Rhythms
Under natural conditions, environmental time cues, such as the onset of light or dark, entrain the free-running rhythm to the light–dark cycle of the environment. In the laboratory it is possible to entrain the circadian rhythms of free-running animals with short pulses of light or dark administered every 24 hours (see the bottom panel of Figure 52.11).
In mammals, the master circadian “clock” consists of two clusters of neurons just above the optic chiasm (the area of the brain where the optic nerves come together). These structures are called the suprachiasmatic nuclei (SCN). If they are destroyed, the animal becomes arrhythmic (loses its circadian rhythm) and is just as likely to eat, drink, sleep, or wake at any time of day.
A notable study by Martin Ralph and his colleagues, then at the University of Virginia, demonstrated that the SCN is the source of circadian rhythms. When the SCNs of adult hamsters with typical 24-hour rhythms were destroyed, the animals became arrhythmic. After several weeks of this arrhythmic behavior, the researchers transplanted SCN tissue from hamster fetuses bred for an atypical (mutant) short-day rhythmicity into the original hamsters’ brains. The experiment produced two remarkable results (Figure 52.12). First, circadian rhythms were restored by the transplanted SCN tissue, demonstrating that the SCN is sufficient to generate circadian rhythms—a unique case of a behavior being restored by a neural transplant. Second, the restored circadian rhythms had the period length of the donor strain, demonstrating that the specific phenotype of the behavior was a property of the donor neural tissue, and thus wholly generated by the SCN.
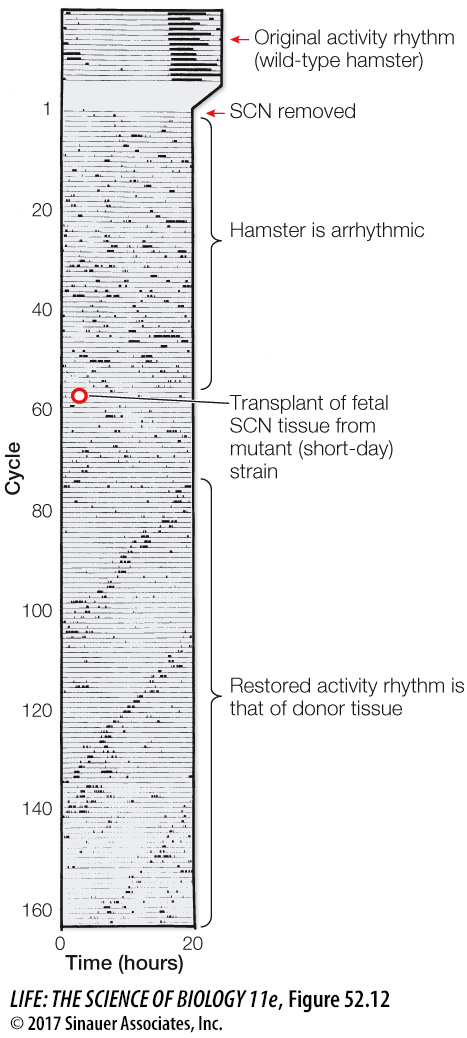
Figure 52.12 The Brain Clock Can Be Transplanted In this experiment, the activity rhythm of a wild-type (i.e., genetically typical) hamster was measured; this animal had a circadian period of 24.5 hours (top of bar). After its SCN was removed (lesioned), the hamster became arrhythmic. SCN tissue from a fetal “short-day” hamster (a mutant strain with a 19-hour circadian period) was then transplanted into the region where the lesion was made in the wild-type hamster. The transplanted tissues restored circadian rhythm in the lesioned hamster, but the restored rhythm had the period of the donor animal.
The molecular mechanism of the circadian clock involves negative feedback loops. Although there are several genes involved, including the per gene discussed in Key Concept 52.2, we can generalize about the mechanism by saying that when certain “clock genes” are expressed in SCN cells, the mRNA enters the cytoplasm, where it is translated. The resulting proteins combine, and the dimer returns to the nucleus as a transcription factor that shuts off the expression of the clock genes until the transcription factor is degraded. The period of this cycle is about a day. These findings show that it is possible to understand circadian rhythms of behavior at all levels, from the molecular rhythm generators to the environmental stimuli that entrain them to the daily cycle of light and dark.
CIRCANNUAL RHYTHMS Seasonal changes in the environment present challenges to many species. Most animals reproduce most successfully if they time their reproductive behavior to coincide with the most favorable time of year for the survival of their offspring. Many species require considerable advance preparation for reproduction. Migratory animals must arrive on their breeding grounds at the right time, and animals that have specialized structures used in mating displays, such as the antlers of deer, moose, and caribou, must grow these structures before the breeding season arrives.
For many species, a change in day length—the *photoperiod—is a reliable indicator of seasonal changes to come. For others, however, change in day length is not a reliable seasonal cue. Hibernators, for example, spend long months in dark burrows underground but must be physiologically prepared to breed almost as soon as they emerge in the spring. A bird overwintering near the equator cannot use changes in photoperiod as a cue to time its migration to its temperate-zone breeding grounds. When held under constant laboratory conditions, such animals show endogenous circannual rhythms that keep track of the time of year. Unlike for circadian rhythms, the neural basis for circannual rhythms is unknown.
*connect the concepts Photoperiodism is also a common phenomenon in plants that have to anticipate seasonal changes, and many do so by being sensitive to the length of the night. See Key Concept 38.2.