Animals use multiple modalities to communicate
As individual animals interact, they exchange information; therefore animal behaviors can evolve into systems of information exchange, or communication. The behaviors of individuals may become elaborated into communication signals, but only if the transmission of information benefits both the sender and the receiver. To understand why these conditions must be met, consider male courtship displays, which can be quite bizarre, time-
Animals communicate using a variety of sensory modalities that vary in the nature of the signal produced, the specificity of the information conveyed, the speed and persistence of the signal, and its suitability in different environments. Behavioral physiologists interested in communication must take into consideration the sensory and motor characteristics of their study animals, the physics of the communication modalities they use, and the environment in which the communication takes place.
CHEMICAL SIGNALS Because of the diversity of their molecular structures, pheromones can communicate very specific, information-
Pheromones are an effective way to exchange species-
1132
VISUAL SIGNALS Visual signals offer the advantage of rapid delivery of information over considerable distances (depending on the environment and the visual acuity of the receiver); they also convey the exact position of the signaler. Signal content can be enhanced by movements (as in a courtship display) or by different postures. Effective visual signals, however, require sufficient light, and the receiver must be looking at the signaler. Thus visual communication is not particularly useful at night or in environments that lack light, such as caves and ocean depths. Some species have overcome this constraint with light-
Another drawback of visual signals is that they can be intercepted by other species. There are predatory firefly species, for example, that mimic the flash pattern of females of other species. A male that approaches the mimicking “female” becomes a meal rather than a mate. Thus deception can be part of animal communication systems, just as it is part of human communication.
ACOUSTIC SIGNALS Sound cannot convey complex information as rapidly as visual signals can. But acoustic signals, unlike visual signals, can be used at night and in dark environments. They are not hindered by objects that would interfere with visual signals, so they can be transmitted in complex environments such as forests. They are often better than visual signals at getting the attention of a receiver because the receiver does not have to be looking at the signaler for the message to be received. Sounds are also useful for communicating over long distances. Even though the intensity of a sound decreases with distance from the source, loud sounds can transmit information over much longer distances than visual signals can. The complex songs of humpback whales, when produced at ocean depths of about 1,000 meters, can be heard hundreds of kilometers away, allowing these whales to locate one another across vast expanses of ocean. Humpback whale populations that feed during the summer in the north Pacific Ocean migrate thousands of kilometers to winter breeding areas in either the eastern Pacific Ocean, the mid-
The information content of acoustic signals can be increased by varying their frequency, as you can see in the sonograms of the species-
1133
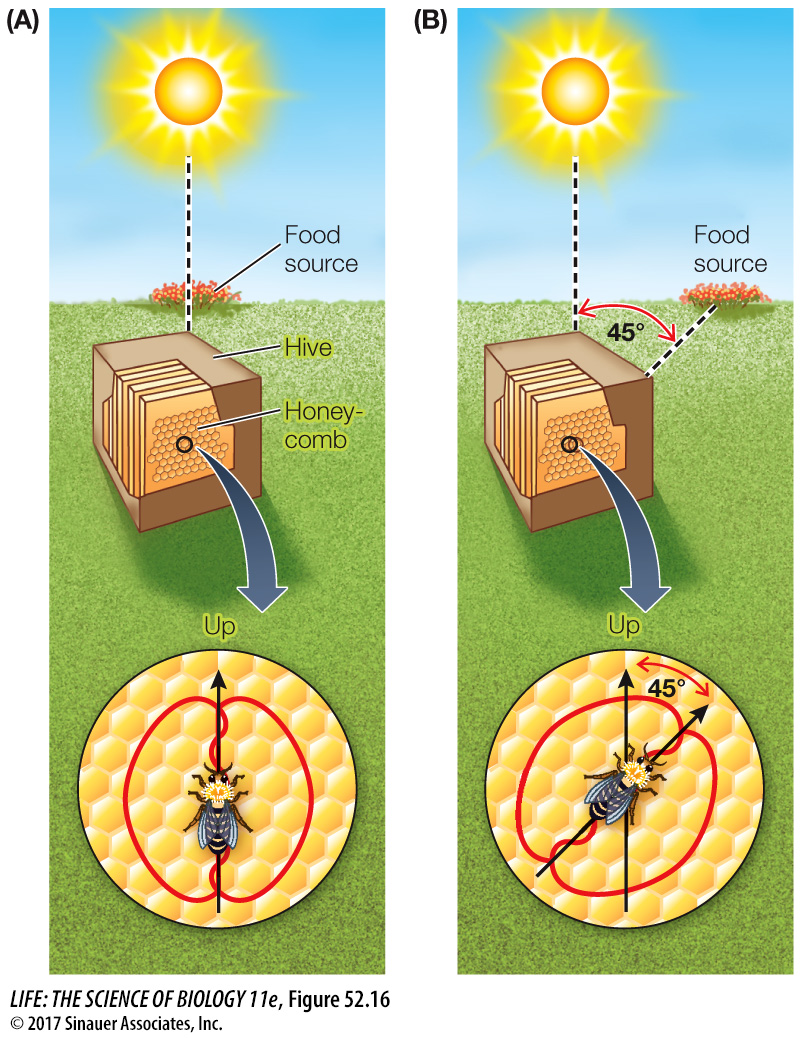
MECHANOSENSORY SIGNALS Animals in close contact with one another can communicate by touch. A classic case of mechanosensory communication is the dance of honey bees (Apis spp.), first described by Karl von Frisch. Honey bees have a spectacular ability to navigate and can accurately communicate the location of food sources as far away as 10 kilometers. When a forager bee finds food, she returns to the hive and communicates her discovery to her hivemates by performing a vigorous waggle dance in the dark hive on the vertical surface of the honeycomb. Other bees follow the dancer and receive her message.
The waggle dance conveys information about both the distance and the direction of the food source. The dancing bee repeatedly traces out a figure-
Activity 52.2 Honey Bee Dance Communication
www.life11e.com/
The distance to the food source is communicated by the duration of the waggle portion of the dance. The farther away the food source is, the longer the duration of each waggle run. When food is close to the hive, the waggle portion of the dance becomes so short that it appeared to von Frisch that it was a different dance, which he called a round dance. Thus the honey bee has a dance language that communicates both the distance and the direction to a food source.
When challenged to prove that the bees were not simply using an odor trail to find the indicated food source, von Frisch responded with a very common sense observation. Bees returning from a new food source fly around barriers such as buildings, but the recruits going out to the food source fly over the barriers in a “beeline,” which would be impossible if they were following an odor trail. Careful observation is still one of the best tools for studying behavior.
COMMUNICATION IN MULTIPLE SENSORY MODALITIES Avoiding ambiguity is a high priority in any signaling system. Signal specificity is enhanced if multiple sensory modalities are used. Courtship behavior in fruit flies, for example, involves visual, tactile, chemical, and acoustic signals (see Figure 52.4A). The male fruit fly orients toward the female’s line of vision (visual signal) and taps her body with his foreleg (tactile signal). Upon detecting pheromones in her cuticle (chemical signal), the male begins to vibrate one wing, producing a species-