Chapter 13
RECAP 13.1
DNA was located in the eukaryotic cell nucleus, where chromosomes carrying genes were located. The amount of DNA was the same in somatic cells of an organism, and halved in the products of meiosis, as expected by the genetic material. Different species had different amounts of DNA, just as they seemingly had different numbers of genes.
Avery and his colleagues performed genetic experiments using cell extracts of one strain of bacteria (A) that could change a recipient strain (B) that was genetically different into the extracted strain (A). When they pretreated the extract of the donor strain with enzymes that hydrolyzed proteins, transformation still occurred. So proteins in the donor extract were not responsible for genetic transformation.
Bacteriophage T2 has only two types of molecules, DNA and protein. So labeling one or the other could indicate which got into a host cell to cause genetic changes.
The proposed experiments might use S strain pneumococcus and transform R strain, as in Figure 13.1. Incubate separate batches of S strain bacteria in 32P or 35S. Make cell-
free extracts of the S strains. Incubate with R cells and look for their transformation to the S phenotype. Then check to see if there is 32P or 35S label in the newly transformed cells. It would be expected that only 32P label (DNA) would enter the cells.
RECAP 13.2
X-
ray diffraction indicated that DNA is double- stranded and twisted into a helix. There were indications that the bases were inside the helix, and the sugars and phosphates on the outside. Base composition data from many organisms showed that the percentages of the purine A = those of the pyrimidine T, and that the percentages of the purine G = those of the pyrimidine C. This suggested that A might be opposite T on the inside of the double helix, and G opposite C. When Watson and Crick built molecular models with the atoms and bonds of polynucleotide strands, the base pairing was confirmed, as the A- T and G- C pairs fit nicely together. The double-
stranded structure is essential in the replication of DNA, as the opposite strands can each act as a template for a new strand, so that two new identical strands are made. This is key in the replication of the genetic material when cells divide. The two strands can unravel at places, exposing the bases in the inside for gene expression. This is important because genes must be expressed for the phenotype. The bases in DNA expose chemical groupings that can interact with groups on proteins. These include polar groups (e.g., C=O) on the bases that can attract oppositely polar groups (e.g., NH2) on proteins, as well as form hydrogen bonds with groups on proteins.
RECAP 13.3
The bacteria were prelabeled with both strands of DNA with heavy 15N. After one round of replication in light 14N, three models for DNA replication had different predictions:
Conservative: There would be original DNA (all heavy) and new DNA (all light) in equal amounts. This did not occur.
Dispersive: There would be light, heavy, and intermediate DNA in no fixed proportions. This did not occur.
Semiconservative: There would be only intermediate DNA, with one light and one heavy strand. This did occur.
The five proteins involved in DNA replication are DNA helicase (unwinds the double helix), single-
strand binding proteins (stabilize and keep apart the two strands in unwound regions), primase (binds to DNA to make a short primer), DNA polymerase (adds nucleotides to a growing chain), and DNA ligase (seals up nicks in DNA, due to its lagging strand replication in short pieces). The special sequences at the ends of chromosomes are called telomeres, and they can be replicated if necessary. After DNA replication, the primer regions at the 3′ ends of the long DNA in chromosomes are removed. This shortens the DNA and makes it unstable. In some cells, such as gamete-
producing cells and cancer cells, the telomeric sequences are recognized by an enzyme complex called telomerase, which catalyzes the replication of any lost telomeric sequences and keeps the DNA at its original length. DNA replication adds new nucleotides to the 3′ end of DNA, where there is an —OH group on the sugar at the 3′ position. If there is no —OH group, there cannot be a condensation reaction and formation of a bond to the next nucleotide, so replication stops.
RECAP 13.4
If there were a mutation in S phase before meiosis (e.g., a T changed to a C), the replicated strand would be mispaired; e.g.,
AAGGTT → AAGGCT TTCCAA TTCCAA If this mispairing was not recognized and repaired, a gamete formed might have the mutation:
AAGGCT
TTCCAA
A-
14 And after the cell containing the mutation divided, one of the daughters would have the mutation and the other would retain the normal sequence.
Top strand replicated: AAGGCT TTAAGA Mutation Bottom strand replicated: AAGGTT TTCCAA Normal If the colon cancer cell had a mutation in a tumor suppressor gene, it might not be repaired. This would lead to unregulated cell division.
RECAP 13.5
Primers in PCR bind to a short region of DNA and allow elongation of that strand of DNA when DNA polymerase is added along with nucleotide.
If a primer has a species-
specific sequence— that is, a sequence of nucleotides complementary to a sequence unique to a certain organism— PCR will amplify a target DNA extracted from a field sample that has the unique sequence. This amplification will show that the organism with that sequence is present.
WORK WITH THE DATA, P. 277
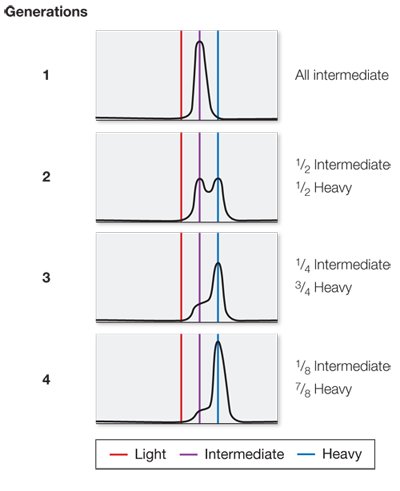
These data fit the semiconservative model of DNA replication because the heavy strands were templates for new light strands; after one round of replication, all the DNA had one heavy (original) strand and one light (newly made) strand, and so was intermediate in weight.
Generation Percent heavy DNA Percent intermediate DNA Percent light DNA 1 0 100 0 2 0 50 50 3 0 25 75 4 0 12.5 87.5 After seven generations there would be about 1.5 percent intermediate DNA and 98.5 percent light DNA.
In the first generation, the bands would be the same as in Figure B: all intermediate. In the second generation, 1/2 would be intermediate and 1/2 heavy. In the third generation, 1/4 would be intermediate and 3/4 heavy. In the fourth generation, 1/8 would be intermediate and 7/8 heavy.
In a conservative model, the first generation would be 1/2 heavy, 1/2 light; the second generation 1/4 heavy, 3/4 light; the third generation 1/8 heavy, 7/8 light; and the fourth generation 1/16 heavy, 15/16 light. In a dispersive model, the first generation would be all intermediate; the second generation all half-
way between intermediate and light; the third generation all half- way between the second generation peak and light; and the fourth generation all half- way between the third generation peak and light. 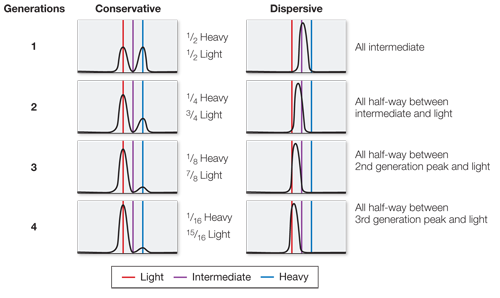
FIGURE QUESTIONS
Figure 13.6 Hydrogen bonds occur between the bases on opposite strands, within base pairs. Covalent bonds occur between the atoms that make up nucleotides and between the nucleotides in a DNA strand. van der Waals forces occur between the flat bases that stack on top of each other within the double helix, stabilizing them in the stacking.
Figure 13.13 On opposite sides of the origin, the leading strand will be the lagging strand, and vice-
Figure 13.17 Shortened telomeres are unstable. Also, without telomerase there is unreplicated DNA. For these two reasons, gamete-
Figure 13.19 Both PCR primers in the test tube and DNA replication primers in the cell act to begin DNA replication. However, PCR primers can be complementary to any DNA strand, whereas DNA replication primers bind only to the origin of replication.
APPLY WHAT YOU’VE LEARNED
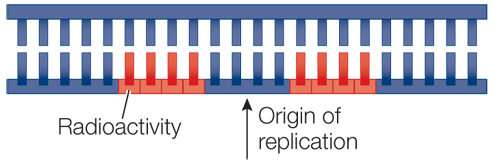
It will take about 280 hours to complete the synthesis of the shortest DNA strand from one starting point. This is much longer than the 8 hours that the cell has to complete DNA replication, so no one can claim that DNA replication involves just one starting point. The results of the calculation suggest that multiple starting points must be involved in replicating a strand of DNA.
The results suggest that multiple starting points for synthesis are present in a single DNA molecule. The number of starting points corresponds to the number of radioactive bands. This result is consistent with the hypothesis made based on the calculations in Question 1.
The results suggest that DNA synthesis is bidirectional. This is shown by the presence of two dark patches running 180 degrees from each starting point of synthesis, followed by lighter patches indicating a period of dilution during the change from radioactive to nonradioactive thymidine. If DNA synthesis were unidirectional, we would see single dark patches followed by lighter patches to either the left or the right but not in both directions.