Mitochondria Contain DNA Located in the Matrix
Although the vast majority of DNA in most eukaryotes is found in the nucleus, some DNA is present within the mitochondria of animals, plants, and fungi and within the chloroplasts of plants. Many lines of evidence indicate that mitochondria and chloroplasts evolved from eubacteria that were engulfed into ancestral cells containing a eukaryotic nucleus, forming endosymbionts (Figure 12-7). Over evolutionary time, most of the bacterial genes were lost from organelle DNA. Some, such as genes encoding proteins involved in nucleotide, lipid, and amino acid biosynthesis, were lost because their functions were provided by genes in the nucleus of the host cell. Other genes encoding components of the present-day organelles were transferred to the nucleus. However, mitochondria and chloroplasts in today’s eukaryotes retain DNA encoding some proteins essential for organelle function as well as the ribosomal and transfer RNAs required for synthesis of those proteins. Thus eukaryotic cells have multiple genetic systems: a predominant nuclear system and secondary systems with their own DNA, ribosomes, and tRNAs in mitochondria and chloroplasts.
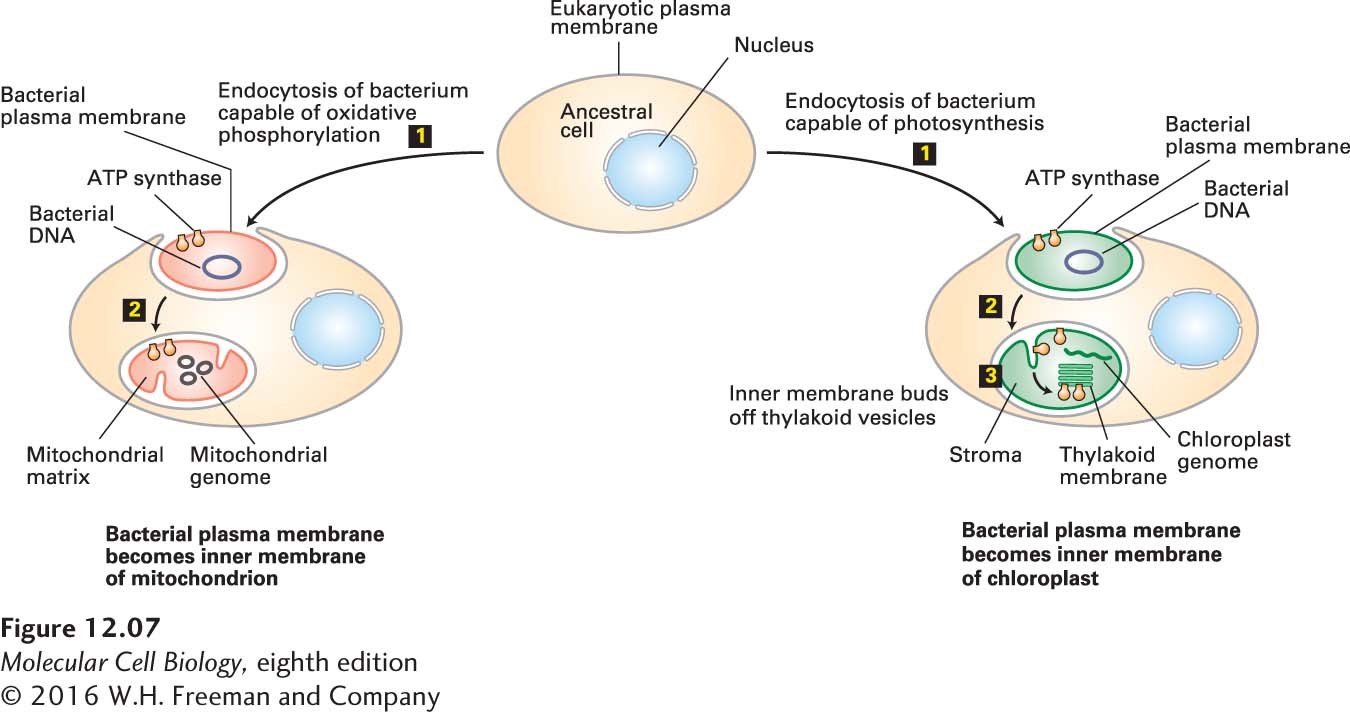
FIGURE 12-7 Endosymbiont hypothesis for the evolutionary origin of mitochondria and chloroplasts. Endocytosis of a bacterium by an ancestral eukaryotic cell (step 1) would generate an organelle with two membranes, the outer membrane derived from the eukaryotic plasma membrane and the inner one from the bacterial membrane (step 2). Proteins localized to the ancestral bacterial membrane would retain their orientation, such that the portion of the protein once facing the extracellular space would now face the intermembrane space. For example, the F1 subunit of ATP synthase, localized to the cytosolic face of the bacterial membrane, would face the matrix of the evolving mitochondrion (left) or chloroplast (right). Budding of vesicles from the inner chloroplast membrane, such as occurs during development of chloroplasts in contemporary plants, would generate the thylakoid membranes with the F1 subunit remaining on the cytosolic face, facing the chloroplast stroma (step 3). The organelle DNAs are indicated. Membrane surfaces facing a shaded area are cytosolic faces; surfaces facing an unshaded area are exoplasmic faces.
The mitochondrial DNA (mtDNA) is located in the mitochondrial matrix (see Figure 12-6). As judged by the number of yellow fluorescent “dots” of mtDNA, a Euglena gracilis cell—a simple, single-celled eukaryote—contains at least 30 mtDNA molecules (Figure 12-8). Replication of mtDNA and division of the mitochondria can be followed in live cells using time-lapse microscopy. Such studies show that in most organisms, mtDNA replicates throughout interphase. At mitosis, each daughter cell receives approximately the same number of mitochondria, but because there is no mechanism for apportioning exactly equal numbers of mitochondria to the daughter cells, some cells contain more mtDNA than others. Thus the total amount of mtDNA in a cell depends on the number of mitochondria, the size of the mtDNA molecules, and the number of mtDNA molecules per mitochondrion. Each of these parameters varies greatly between cell types. In a typical human cell, there are about 1000–2000 mtDNA molecules per nucleus; however, a human egg has about 500,000 mtDNA molecules, and a sperm has only about 100.
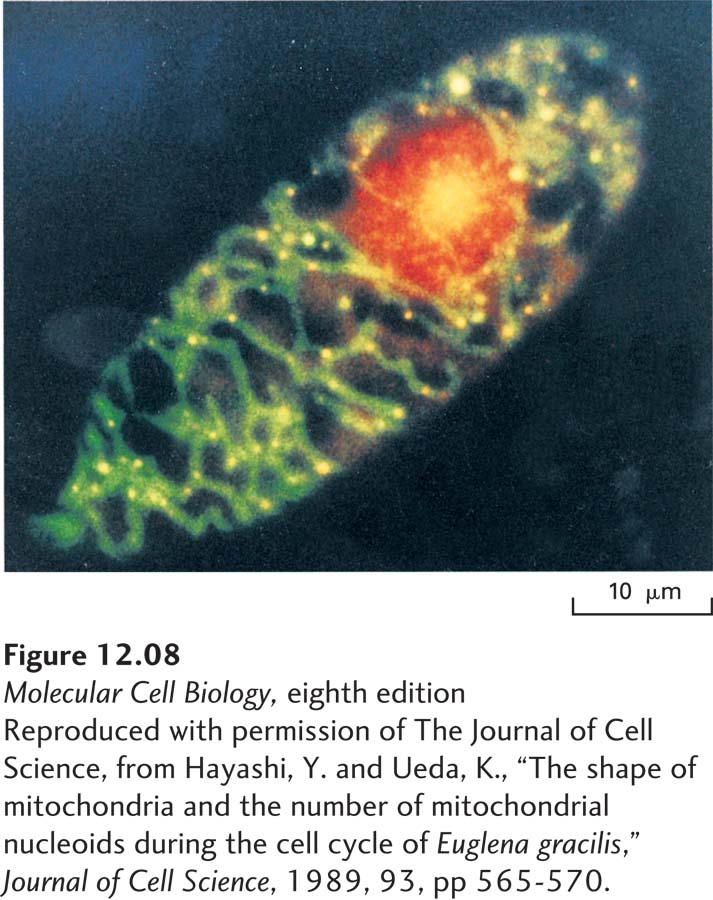
EXPERIMENTAL FIGURE 12-8 Dual staining reveals the multiple mitochondrial DNA molecules in a growing Euglena gracilis cell. Cells were treated with a mixture of two dyes: ethidium bromide, which binds to DNA and emits a red fluorescence, and DiOC6, which is incorporated specifically into mitochondria and emits a green fluorescence. Thus the nucleus emits a red fluorescence, and areas rich in mitochondrial DNA fluoresce yellow—a combination of red DNA and green mitochondrial fluorescence.
[Reproduced with permission of The Journal of Cell Science, from Hayashi, Y. and Ueda, K., “The shape of mitochondria and the number of mitochondrial nucleoids during the cell cycle of Euglena gracilis,” Journal of Cell Science, 1989, 93, pp 565-570.]
Studies of mutants in yeasts and other single-celled organisms first indicated that mitochondria exhibit cytoplasmic inheritance (Figure 12-9). For instance, yeast cells with the petite mutation exhibit structurally abnormal mitochondria and are incapable of oxidative phosphorylation. As a result, petite cells grow more slowly than wild-type cells and form smaller colonies. Genetic crosses between different (haploid) yeast strains showed that the petite mutation does not segregate with any known nuclear gene or chromosome. In later studies, most petite mutants were found to contain deletions of mtDNA.
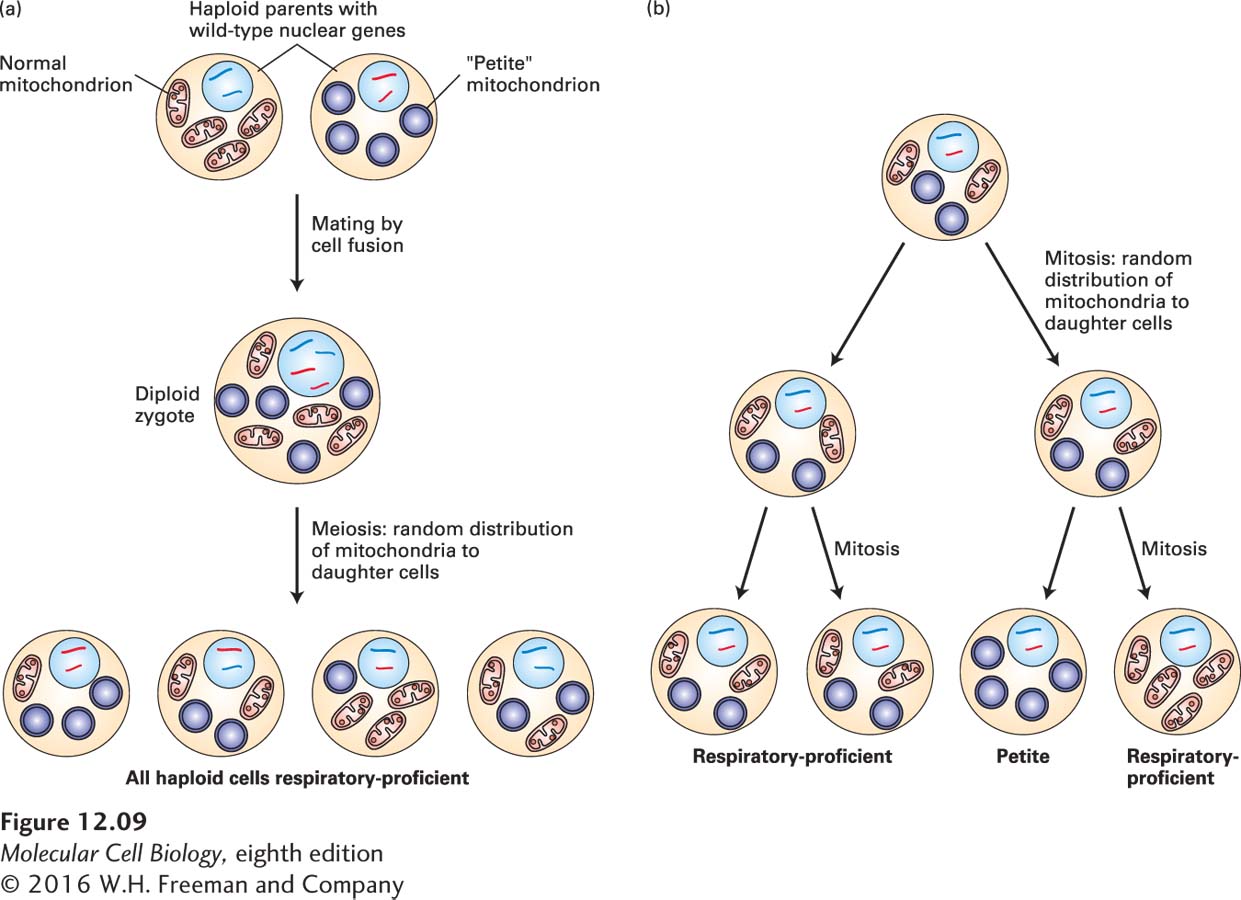
FIGURE 12-9 Cytoplasmic inheritance of an mtDNA petite mutation in yeast. Petite-strain mitochondria are defective in oxidative phosphorylation owing to a deletion in mtDNA. (a) Haploid yeast cells fuse to produce a diploid cell that undergoes meiosis, during which random segregation of parental chromosomes and mitochondria containing mtDNA occurs. Note that alleles for genes in nuclear DNA (represented by large and small nuclear chromosomes colored red and blue) segregate 2:2 during meiosis (see Figure 6-5). In contrast, since yeast normally contain some 50 mtDNA molecules per cell, most products of meiosis contain both normal and petite mtDNAs and are capable of respiration. (b) As these haploid cells grow and divide mitotically, the cytoplasm (including the mitochondria) is randomly distributed to the daughter cells. Occasionally, a cell is generated that contains only petite mtDNA and yields a petite colony. Thus formation of such petite cells is independent of any nuclear genetic marker.
In the mating by fusion of haploid yeast cells, both parents contribute equally to the cytoplasm of the resulting diploid; thus inheritance of mitochondria is biparental (see Figure 12-9a). In mammals and most other multicellular organisms, however, the sperm contributes little (if any) cytoplasm to the zygote, and virtually all the mitochondria in the embryo are derived from those in the egg, not the sperm. Studies in mice have shown that 99.99 percent of mtDNA is maternally inherited, but a small part (0.01 percent) is inherited from the male parent. In higher plants, mtDNA is inherited exclusively in a uniparental fashion through the female parent (egg), not the male (pollen).