Three of the Four Stages in Photosynthesis Occur Only During Illumination
The photosynthetic process in plants can be divided into four stages (see Figure 12-38), each localized to a defined area of the chloroplast: (1) absorption of light, generation of high-energy electrons, and formation of O2 from H2O; (2) electron transport leading to reduction of NADP+ to NADPH, and generation of a proton-motive force; (3) synthesis of ATP; and (4) conversion of CO2 into carbohydrates, commonly referred to as carbon fixation. The enzymes that incorporate CO2 into chemical intermediates and then convert them to starch are soluble constituents of the chloroplast stroma; the enzymes that form sucrose from three-carbon intermediates are in the cytosol. All four stages of photosynthesis are tightly coupled and controlled so as to produce the amount of carbohydrate required by the plant. All the reactions in stages 1–3 are catalyzed by multiprotein complexes in the thylakoid membrane. The generation of a proton-motive force and the use of that proton-motive force to synthesize ATP resemble stages III and IV of mitochondrial oxidative phosphorylation.
Stage 1: Absorption of Light Energy, Generation of High-Energy Electrons, and O2 Formation The initial step in photosynthesis is the absorption of light by chlorophylls attached to proteins in the thylakoid membranes. Like the heme component of cytochromes, chlorophylls consist of a porphyrin ring attached to a long hydrocarbon side chain (Figure 12-39). In contrast to the hemes (see Figure 12-20), chlorophylls contain a central Mg2+ ion (rather than Fe2+) and have an additional five-member ring. The energy of the absorbed light is ultimately used to remove electrons from a donor (water in the case of green plants), forming oxygen:
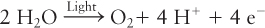
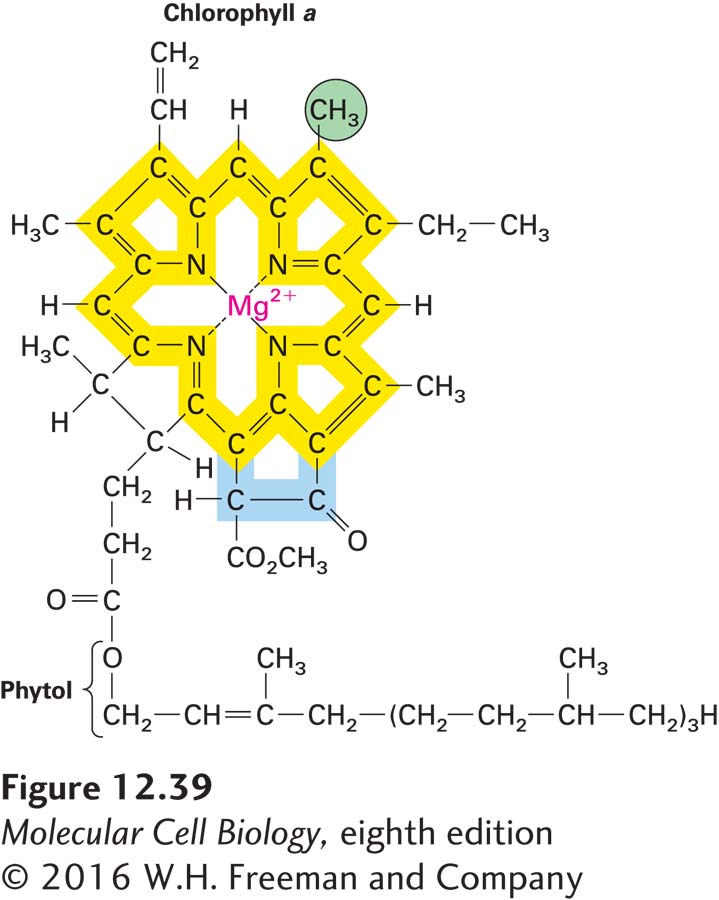
FIGURE 12-39 Structure of chlorophyll a, the principal pigment that traps light energy. Electrons are delocalized among three of chlorophyll a’s four central rings (yellow) and the atoms that interconnect them. In chlorophyll, a Mg2+ ion, rather than the Fe2+ ion found in heme, sits at the center of the porphyrin ring, and an additional five-member ring (blue) is present; otherwise, the structure of chlorophyll is similar to that of heme, found in molecules such as hemoglobin and cytochromes (see Figure 12-20a). The hydrocarbon phytol “tail” facilitates the binding of chlorophyll to hydrophobic regions of chlorophyll-binding proteins. The CH3 group (green) is replaced by a formaldehyde (CHO) group in chlorophyll b.
The electrons are transferred to a primary electron acceptor, a quinone designated Q, which is similar to CoQ in mitochondria. In plants, the oxidation of water takes place in a multiprotein complex called photosystem II (PSII).
Quantum mechanics established that light, a form of electromagnetic radiation, has properties of both waves and particles. When light interacts with matter, it behaves as discrete packets of energy (quanta) called photons. The energy of a photon is proportional to the frequency of the light wave, and thus inversely proportional to its wavelength. Thus photons of shorter wavelengths have higher energies. The energy of visible light is considerable. Light with a wavelength of 550 nm (550 × 10−7 cm), typical of sunlight, has about 52 kcal of energy per mole of photons. This is enough energy to synthesize several moles of ATP from ADP and Pi if all the energy were used for this purpose.
Stage 2: Electron Transport and Generation of a Proton-Motive Force Electrons move from the quinone primary electron acceptor through a series of electron carriers until they reach the ultimate electron acceptor, usually the oxidized form of nicotinamide adenine dinucleotide phosphate (NADP+), reducing it to NADPH. The structure of NADP+ is identical to that of NAD+ except for the presence of an additional phosphate group. Both molecules gain and lose electrons in the same way (see Figure 2-33). In plants, the reduction of NADP+ takes place in a complex called photosystem I (PSI) (Figure 12-38). The transport of electrons in the thylakoid membrane is coupled to the movement of protons from the stroma to the thylakoid lumen, forming a pH gradient across the membrane (pHlumen < pHstroma). This process is analogous to the generation of a proton-motive force across the inner mitochondrial membrane and in bacterial membranes during electron transport (see Figure 12-29).
Thus the overall reaction of stages 1 and 2 can be summarized as
Stage 3: Synthesis of ATP Protons move down their concentration gradient from the thylakoid lumen to the stroma through the chloroplast F0F1 complex (ATP synthase), which couples proton movement to the synthesis of ATP from ADP and Pi, as we have seen for the ATP synthases in mitochondria and bacteria (see Figures 12-31, 12-32, and 12-34).
Stage 4: Carbon Fixation The NADPH and ATP generated by stages 2 and 3 of photosynthesis provide the energy and the electrons to drive the synthesis of polymers of six-carbon sugars from CO2 and H2O. The overall chemical equation is written as
6 CO2 + 18 ATP4− + 12 NADPH + 12 H2O → C6H12O6 + 18 ADP3− + 18 Pi2− + 12 NADP+ + 6 H+
The reactions that generate the ATP and NADPH used in carbon fixation are directly dependent on light energy; thus stages 1–3 are called the light reactions of photosynthesis. The reactions in stage 4 are indirectly dependent on light energy; they are sometimes called the “dark” reactions of photosynthesis because they can occur in the dark, using the supplies of ATP and NADPH generated by light energy (see Figure 12-38). However, the reactions in stage 4 are not confined to the dark; in fact, they occur primarily during illumination.