Cell Migration Involves the Coordinate Regulation of Cdc42, Rac, and Rho
How does each of these small GTP-binding proteins contribute to the regulation of cell migration? To answer this question, researchers developed an in vitro wound-healing assay (Figure 17-44a). Cells in culture are grown in a petri dish with growth factors until they are confluent and form a tight monolayer, at which point they stop dividing. The cell monolayer is then scratched with a needle to remove a swath of cells, generating a “wound” containing a free edge of cells. The cells on the edge sense the loss of their neighbors and, in response to components of the extracellular matrix now exposed on the dish surface, move to fill up the empty wound area (Figure 17-44b). To do this, they orient themselves toward the empty area, first extending a lamellipodium and then moving in that direction. In this way, one can study the induction of directed cell migration in vitro.
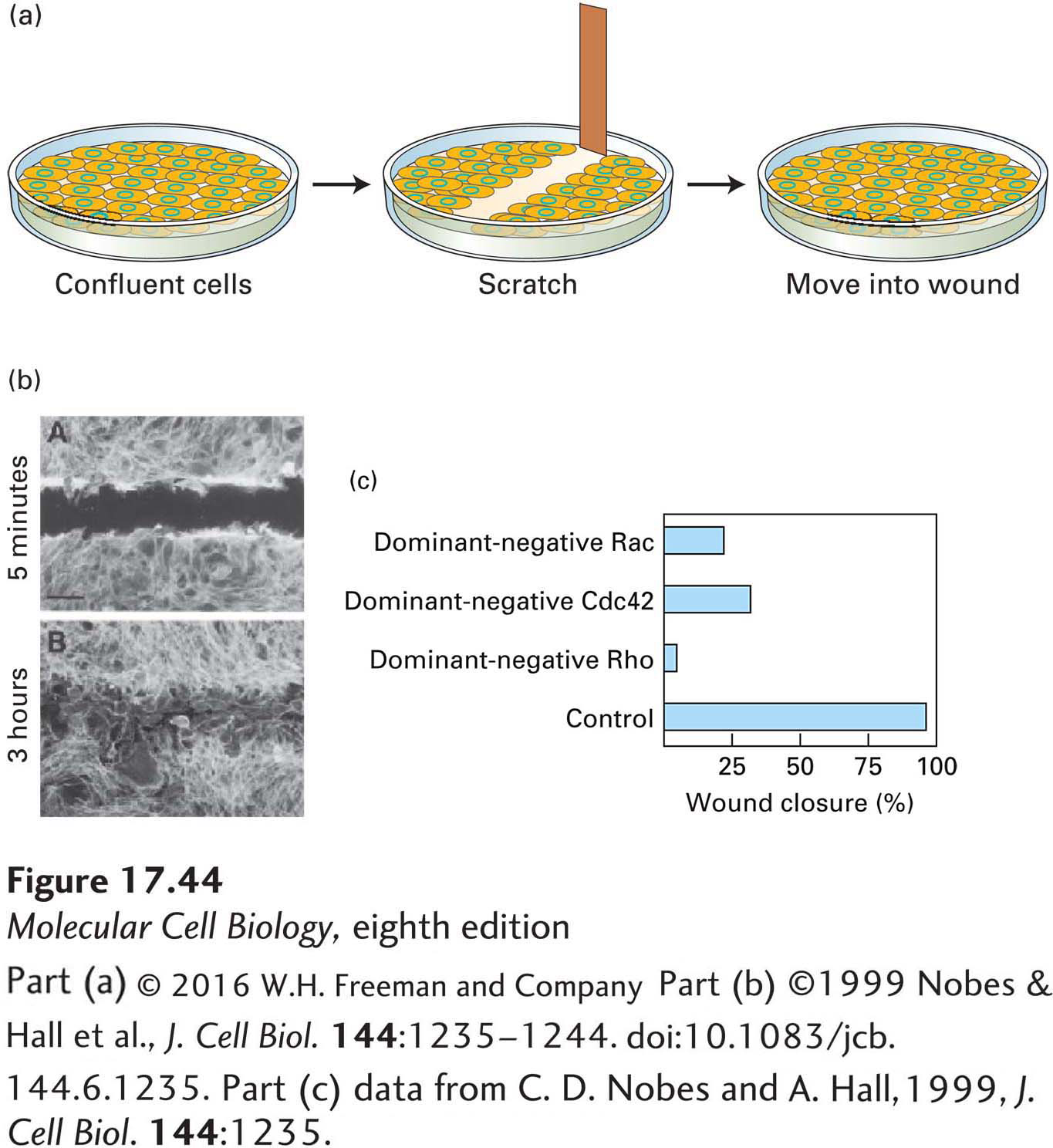
EXPERIMENTAL FIGURE 17-44 The wounded-cell monolayer assay can be used to dissect signaling pathways in directed cell movement. (a) A confluent layer of cells is scratched to remove a swath about three cells wide to generate a free cell border. The remaining cells detect the free space and newly exposed extracellular matrix and, over a period of hours, fill in the area. (b) Localization of actin in a wounded monolayer 5 minutes and 3 hours after scratching; after 3 hours, cells have migrated into the wounded area. (c) Effect of introducing dominant-negative Cdc42, Rac, and Rho into cells at the wound edge; all affect wound closure.
[Part (b) ©1999 Nobes & Hall et al., J. Cell Biol. 144:1235–1244. doi:10.1083/jcb.144.6.1235. Part (c) data from C. D. Nobes and A. Hall, 1999, J. Cell Biol. 144:1235.]
Using this wounded-cell monolayer assay, researchers have introduced dominant-negative Rac into cells on the wound edge to see how it affects the ability of the cells to migrate and fill the wound. Since Rac is needed for activation of the Arp2/3 complex to form the lamellipodium, it is not surprising that the cells fail to form this structure and do not migrate, and so the wound does not close (Figure 17-44c). A very interesting result is obtained when dominant-negative Cdc42 is introduced into the cells at the wound edge: they can form a lamellipodium, but they do not orient in the correct direction—in fact, they try to migrate in random directions. This observation suggests that Cdc42 is critical for regulating the overall polarity of the cell. Studies from yeast (in which Cdc42 was first described), wounded-cell monolayers, epithelial cells, and neurons reveal that Cdc42 is a master regulator of polarity in many different systems. Part of this regulation in animals involves the binding of Cdc42 to its effector, Par6, a polarity protein that functions in nematodes (in which it was first discovered), neurons, and epithelial cells. We explore these polarity pathways in more detail in Chapter 21.
Studies such as these suggest a general model of how cell migration is controlled (Figure 17-45). Signals from the environment are transmitted to Cdc42, which orients the cell. The oriented cell has high Rac activity at the front, which induces the formation of the leading edge; Rho activity is high in the rear, inducing the assembly of contractile structures and activating the myosin-II-based contractile machinery. It is important to notice that different regions of the cell can have different levels of active Cdc42, Rac, or Rho, so these regulators are controlled locally within the cell. Part of this spatial regulation occurs because some small G proteins can work antagonistically. For example, active Rho can stimulate pathways that lead to the inactivation of Rac. This process might help ensure that no leading-edge structures form at the rear of the cell.
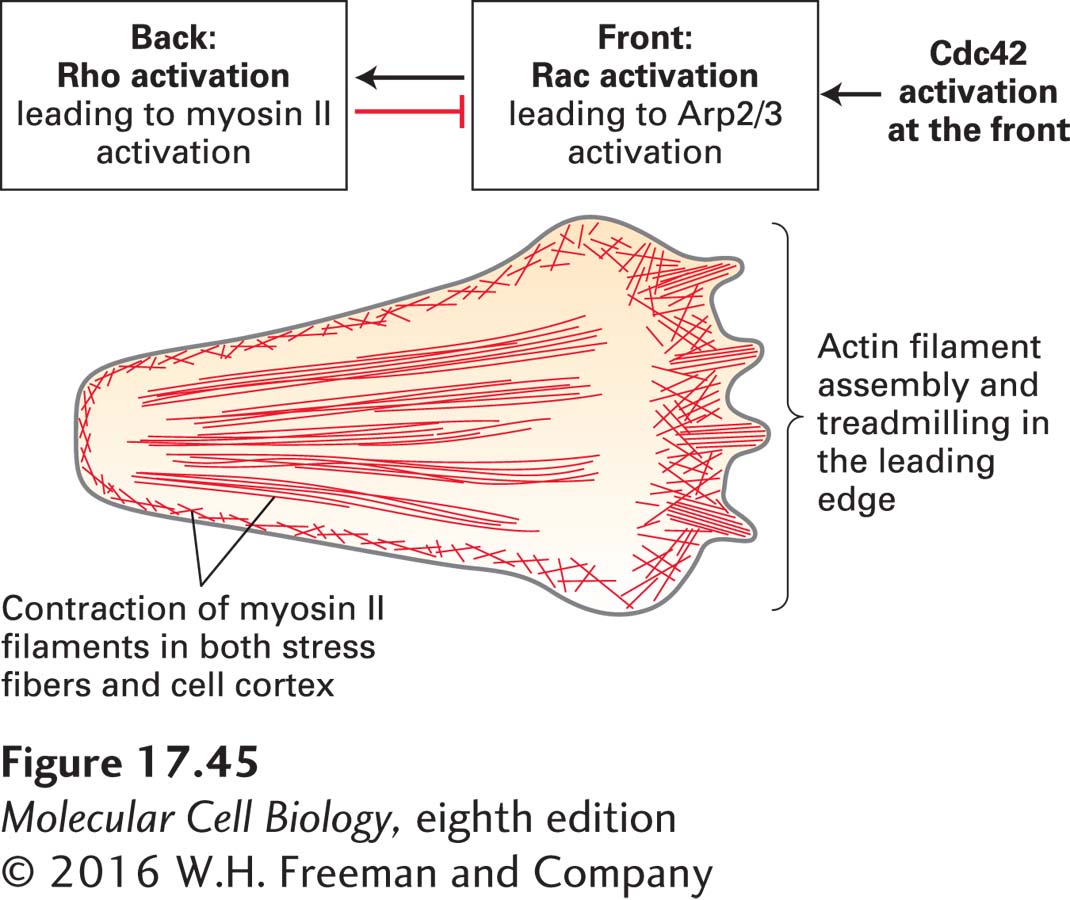
FIGURE 17-45 Contributions of Cdc42, Rac, and Rho to cell movement. The overall polarity of a migrating cell is controlled by Cdc42, which is activated at the front of a cell. Cdc42 activation leads to active Rac in the front of the cell, which generates the leading edge, and active Rho at the back of the cell, which causes myosin II activation and contraction. Active Rho inhibits Rac activation, ensuring the asymmetry of the two active G-proteins.