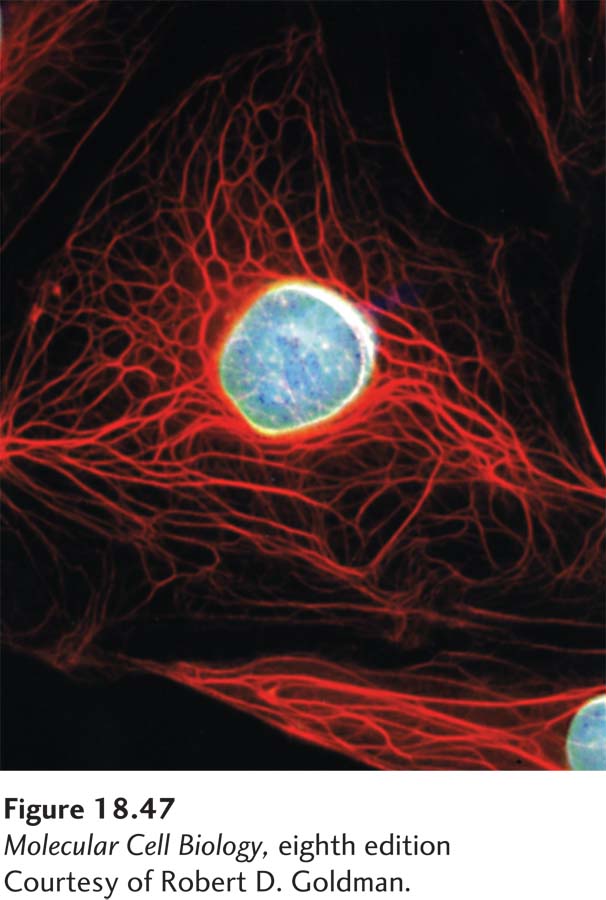
EXPERIMENTAL FIGURE 18-47 Localization of two types of intermediate filaments in an epithelial cell. Immunofluorescence micrograph of an epithelial cell doubly stained with antibodies to keratin (red) and lamin (blue). A meshwork of lamin intermediate filaments can be seen underlying the nuclear membrane, whereas the keratin filaments extend from the nucleus to the plasma membrane.
[Courtesy of Robert D. Goldman.]
The third major filament system of eukaryotes comprises intermediate filaments (Ifs). This name reflects their diameter of about 10 nm, which is intermediate between the 6–8-nm diameter of microfilaments and the myosin thick filaments of skeletal muscle. Intermediate filaments extend throughout the cytoplasm as well as lining the inner nuclear envelope of interphase animal cells (Figure 18-47). Intermediate filaments have several unique properties that distinguish them from microfilaments and microtubules. First, they are biochemically much more heterogeneous—that is, many different, but evolutionarily related, IF subunits exist—and are often expressed in a tissue-dependent manner. Second, they have great tensile strength, as is clearly demonstrated by hair and nails, which consist primarily of the intermediate filaments of dead cells. Third, they do not have an intrinsic polarity like microfilaments and microtubules, and their constituent subunits do not bind a nucleotide. Fourth, because they have no intrinsic polarity, it is not surprising that there are no known motors that use them as tracks. Fifth, although they are dynamic in terms of subunit exchange, they are much more stable than microfilaments and microtubules because the exchange rate is much slower. Indeed, a standard way to purify intermediate filaments is to subject cells to harsh extraction conditions in a detergent so that all membranes, microfilaments, and microtubules are solubilized, leaving a residue that is almost exclusively intermediate filaments. Finally, intermediate filaments are not found in all eukaryotes. Fungi and plants do not have intermediate filaments, and insects have only one class, represented by two genes that express lamins A/C and B.
These properties make intermediate filaments unique and important structures of metazoans. Their importance is underscored by the identification of hundreds of clinical disorders, some of which are discussed here, associated with mutations in genes encoding IF proteins. To understand their contributions to cell and tissue structure, we first examine the structure of IF proteins and see how they assemble into filaments. Next we discuss their dynamics, then describe the different classes of intermediate filaments and the functions they perform.