In Chapter 17, we discussed how the polarity of a migrating cell is regulated by Cdc42, which results in the formation of an actin-based leading edge at the front of the cell and contraction at the back (see Figure 17-45 and Figure 18-53, step 1). It turns out that Cdc42 activation at the cell front also leads to polarization of the microtubule cytoskeleton. This phenomenon was originally studied in wound-healing assays (see Figure 17-44), in which it was noticed that when the cells at the edge of a scratch are induced to polarize and move to fill in the empty space, the Golgi complex is moved to the front of the nucleus toward the cell front. Golgi localization at the front of the cell indicates that the centrosome moves to lie in front of the nucleus (recall that Golgi localization is dependent on the location of the MTOC; see Figures 18-1c, 18-28). Recent studies have suggested how this happens. Active Cdc42 at the front of the cell binds the polarity factor Par6, which results in the recruitment of the dynein-dynactin complex (Figure 18-53, step 2). Cortically localized dynein-dynactin then interacts with microtubules, pulling on them to orient the centrosome and hence the whole radial array of microtubules (Figure 18-53, step 3). This reorientation of the microtubule system leads to the reorganization of the secretory pathway to deliver secretory products, especially integrins to bind the extracellular matrix, to the front of the cell for attachment to the substratum for cell migration (Figure 18-53, step 4).
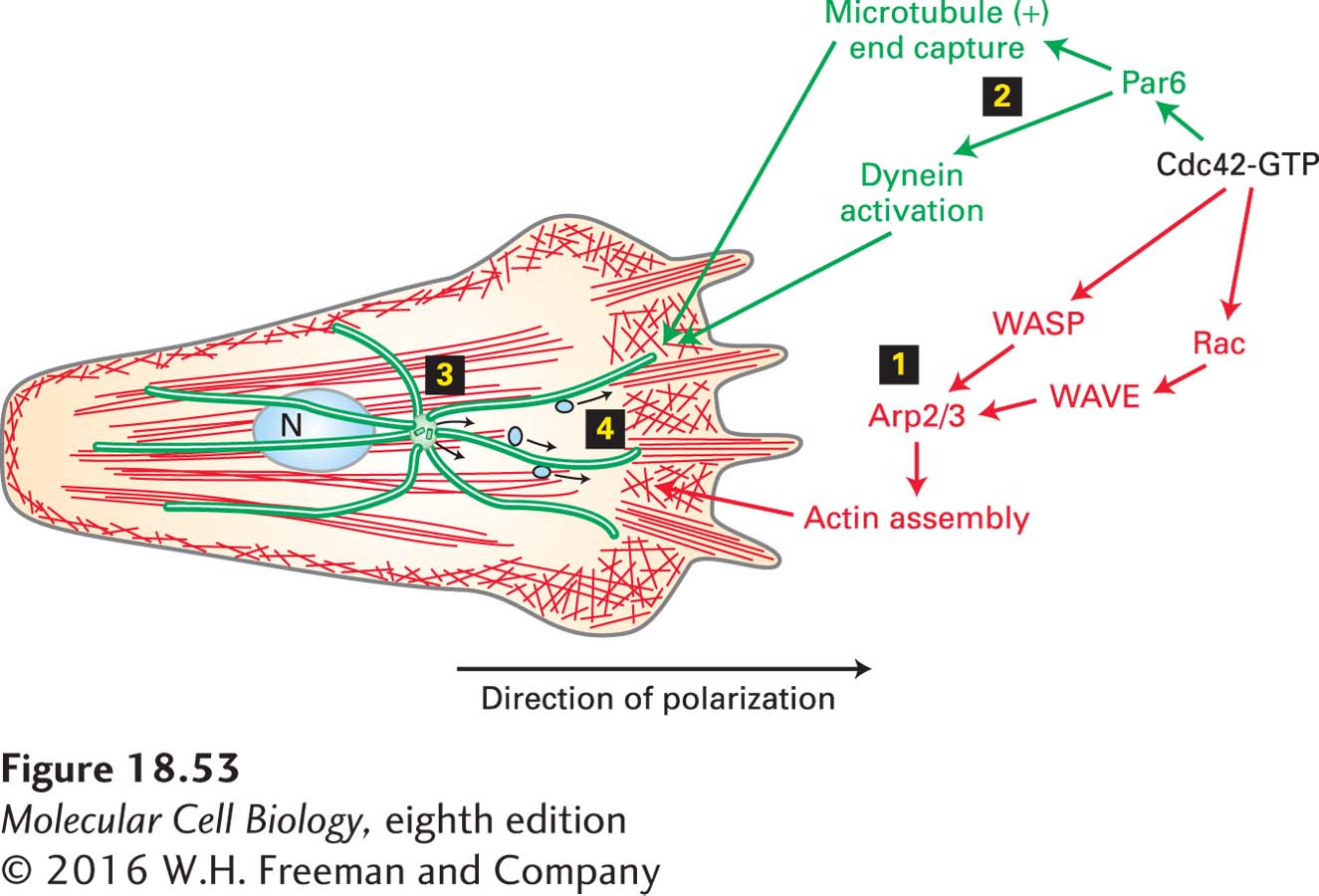
FIGURE 18-53 Independent Cdc42 regulation of microfilaments and microtubules to polarize a migrating cell. Active Cdc42·GTP at the front of the cell leads to Rac and WASP activation, which results in the assembly of a microfilament-based leading edge (step 1). Independently, Cdc42·GTP also leads to the capture of microtubule (+) ends and the activation of dynein (step 2). Dynein pulls on microtubules to orient the centrosome (step 3) toward the front of the cell. This reorientation reorganizes the secretory pathway for the delivery of adhesion molecules carried in secretory vesicles along microtubules to the front of the cell (step 4). See S. Etienne-Manneville et al., 2005, J. Cell Biol. 170:895–901.