The Inner Cell Mass Is the Source of ES Cells
Embryonic stem cells can be isolated from the inner cell mass of early mammalian embryos and grown indefinitely in culture when attached to a feeder-cell layer that provides certain essential growth factors (Figure 21-5a). As mentioned in the chapter introduction, cultured ES cells are pluripotent: they can differentiate into a wide range of cell types of the three primary germ layers, either in culture or after reinsertion into a host embryo. More specifically, mouse ES cells can be injected into the blastocoel of an early mouse embryo and the cell aggregate surgically transplanted into the uterus of a pseudopregnant female. The injected ES cells will participate in forming most, if not all, tissues of the resultant chimeric mice (see Figure 6-38). Furthermore, the injected ES cells will often give rise to functional sperm and eggs that, in turn, can generate normal live mice.
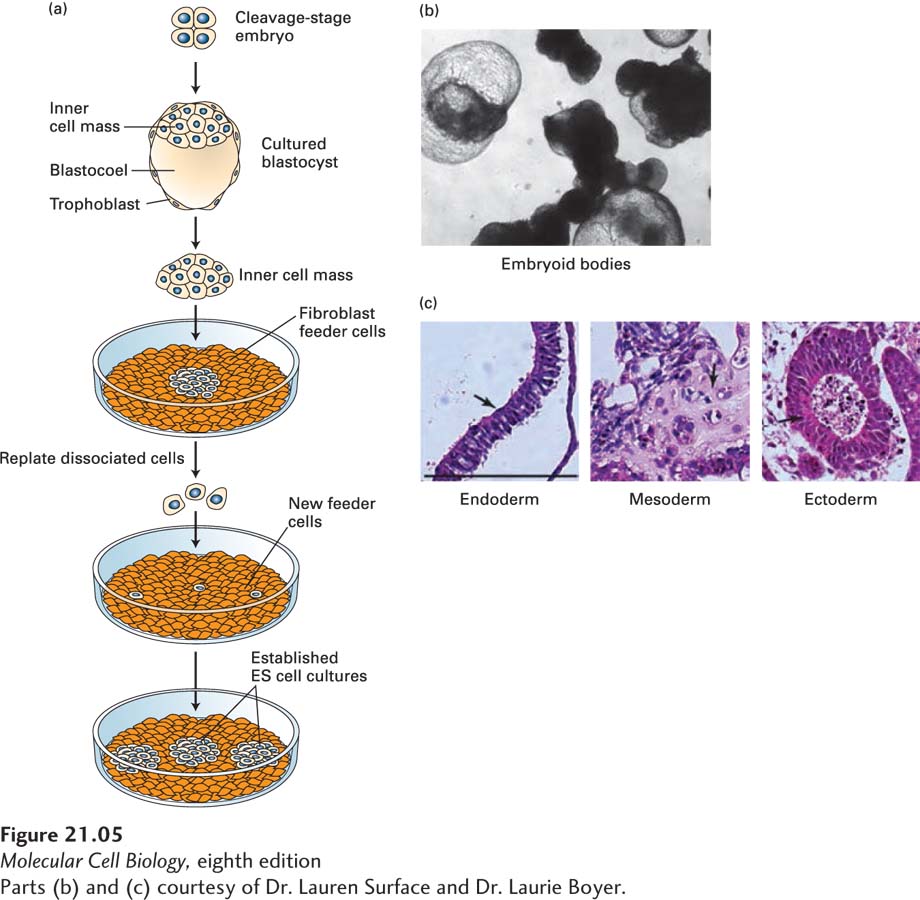
EXPERIMENTAL FIGURE 21-5 Embryonic stem (ES) cells can be maintained in culture and can form differentiated cell types. (a) Human or mouse blastocysts are grown from cleavage-stage embryos produced by in vitro fertilization. The ICM is separated from the surrounding extraembryonic tissues and plated onto a layer of fibroblast cells, which help to nourish the embryonic cells by providing specific protein hormones. When individual cells are replated, they form colonies of ES cells, which can be maintained for many generations and can be stored frozen. ES cells can also be cultured without a fibroblast feeder layer if specific cytokines are added; leukemia inhibitory factor (LIF), for instance, supports growth of mouse ES cells by triggering activation of the Stat3 transcription factor; see J. S. Odorico et al., 2001, Stem Cells 19:193. (b) Embryonic stem cells allowed to differentiate in suspension culture become multicellular aggregates termed embryoid bodies. (c) Hematoxylin- and eosin-stained sections of embryoid bodies that contain derivatives of all three germ layers that are formed from the ICM during embryogenesis. Arrows in the images point to the following tissue types: (left) gut epithelium (endoderm), (middle) cartilage (mesoderm), and (right) neuroepithelial rosettes (ectoderm). Black bar = 100 µm.
[Parts (b) and (c) courtesy of Dr. Lauren Surface and Dr. Laurie Boyer.]
In a more recent variation on these experiments, the host blastocyst is treated with drugs that transiently block mitosis so that its cells become tetraploid (with four copies of each chromosome, incapable of forming differentiated cells and tissues), in contrast to the diploid ES cells that are injected into the blastocyst. In this case, all the cells in the live mice that are born after transplantation of the blastocyst aggregate derive from the donor ES cells. This finding is powerful evidence that single mouse ES cells are indeed pluripotent. Because ethical considerations and, in many countries, legal restrictions preclude similar transplantation experiments with human ES cells, formal proof that they are pluripotent is lacking.
Importantly, both human and mouse ES cells can differentiate into a wide range of cell types in culture. When cultured in suspension, ES cells form multicellular aggregates, called embryoid bodies (Figure 21-5b), that resemble early embryos in the variety of tissues they form. When embryoid bodies are subsequently treated with various combinations of growth factors or transferred to a solid surface, they produce a variety of differentiated cell types, including gut epithelia, cartilage, and neural cells (Figure 21-5c). Under other conditions, ES cells have been induced to differentiate in culture into precursors for various specific cell types, including blood cells and pigmented epithelia; for this reason, ES cells have proved extremely useful in identifying the factors that commit a pluripotent cell to differentiating down a particular cell lineage.
What properties give these cells of the early embryo their remarkable plasticity? As we’ll see in the next section, a variety of actors play a role: DNA methylation, transcription factors, chromatin regulators, and micro-RNAs all affect which genes become active.