Voltage-Sensing S4 α Helices Move in Response to Membrane Depolarization
Our understanding of channel-protein biochemistry is advancing rapidly owing to newly obtained crystal structures for bacterial and Shaker potassium channels and other channels. Transmembrane proteins are notoriously difficult to produce and crystallize, posing unique challenges for the scientist. One method used to obtain crystals of these difficult membrane proteins was to surround them with bound fragments of monoclonal antibodies [F(ab)’s; Chapter 23]; in other cases they were crystallized in complexes with normal protein-binding partners. In both cases the presence of these water-soluble proteins in the complex somehow enhanced crystal formation.
The structures of the channels reveal remarkable arrangements of the voltage-sensing domains, and suggest how parts of the protein move in order to open the channel. As already noted, the K+-channel tetramer, like the Na+-channel monomer, has a pore whose walls are formed by helices S5 and S6 (Figure 22-13a and Figure 22-14). Outside that core structure four arms, or “paddles,” each containing helices S1–S4, protrude into the surrounding membrane and also interact with the outer sides of the S5 and S6 helices; these are the voltage sensors, and they are in minimal contact with the pore. Sensitive electrical measurements suggested that the opening of a voltage-gated Na+ or K+ channel is accompanied by the movement of 12 to 14 protein-bound positive charges from the cytosolic to the exoplasmic surface of the membrane. The moving parts of the protein are the rigid complexes composed of helices S1–S4; S4 accounts for much of the positive charge and is the primary voltage sensor, with a positively charged lysine or arginine every third or fourth residue (Figure 22-14d). Arginines in S4 have been measured moving as much as 1.5 nm as the channel opens, which can be compared with the ~5-nm thickness of the membrane or the 1.2-nm diameter of the α helix itself.
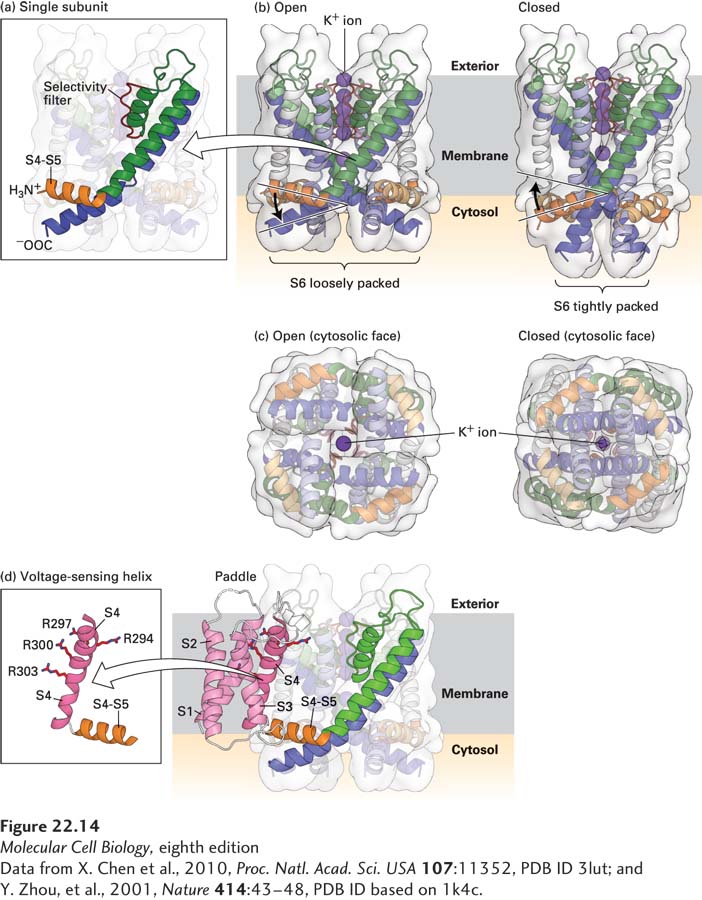
FIGURE 22-14 Molecular structure of a voltage-sensitive K+ channel. Models of the potassium channel single subunit (a) and tetramer (b) as viewed from the side, in open and closed states. The four green (S5) and blue (S6) α helices span the membrane, with the interior of the cell at the bottom and exterior at the top. Note how the helices are tightly packed at the bottom in the closed conformation, so that the K+ ion cannot pass through. (Compare the distances between S5 helices as shown by the curly brackets below (a) and (b).) The S4–S5 linker (orange), located in the cytoplasm, connects the S4 helix (not shown) to the S5 helix. For clarity, helices S1 through S4 have been omitted from the model; they would normally be attached to the end of the S4–S5 linker and protrude from the molecule. (c) Ribbon diagrams of the open and closed states of the channel as viewed from the cytoplasmic face of the membrane. In the open, but not in the closed, state, potassium ions (dark purple) can pass through the pore. (d) Three-dimensional structure of the voltage-sensing “paddles” comprising helixes S1–S4, with the four voltage-sensing arginine (R) residues in S4. These paddles move from near the interior to the exterior of the membrane in response to depolarization. Since each one is attached to an S4–S5 linker, each linker and its attached S5 helix is moved, in turn moving S6 helices, which opens the pore. Note that as shown in (b), the linker between S4 and S5 is pointed upward toward the exoplasmic (exterior) surface in the open channel, pulled upward by the outward movement of the S1–S4 paddles; in contrast the S4–S5 linker is pointing downward in the closed channel when the S1–S4 paddles are nearer the cytosolic surface.
[Data from X. Chen et al., 2010, Proc. Natl. Acad. Sci. USA 107:11352, PDB ID 3lut; and Y. Zhou, et al., 2001, Nature 414:43–48, PDB ID based on 1k4c.]
In the resting state, the positive charges on the S1–S4 complexes (the “paddles”) are attracted to the negative charges on the cytosolic face of the membrane. In the depolarized membrane, these same positive charges become attracted to the negative charges on the exoplasmic (outer) surface of the membrane, causing the S1–S4 paddles to move partly across the membrane—from the cytosolic to the exoplasmic surface. This movement is depicted schematically for the voltage-gated Na+ channel in Figure 22-10 and triggers a conformational change in the protein that opens the channel.
The most unusual aspect of the voltage-sensitive channel structures is the presence of charged groups, for example, arginines, in contact with lipid. The location of the voltage sensor helps to explain earlier experiments in which a non-voltage-sensitive channel was converted into a voltage-sensing channel by adding voltage-sensing domains. Such a result would seem unlikely if the voltage sensors had to be deeply embedded in the core structure.
Studies with mutant Shaker K+ channels support the importance of the S4 helix in voltage sensing. When one or more arginine or lysine residues in the S4 helix of the Shaker K+ channel were replaced with neutral or acidic residues, fewer positive charges than normal moved across the membrane in response to a membrane depolarization, indicating that arginine and lysine residues in the S4 helix do indeed move across the membrane. The structure of the open form of a mammalian voltage-gated K+ channel has been contrasted with the closed structure of a different K+ channel. The results suggest a model for the opening and closing of the channel in response to movements of the voltage sensors across the membrane (see Figure 22-14a, b). In the model, the voltage sensors, composed of helices S1–S4, move in response to voltage and exert a torque on a linker helix that connects S4 to S5:
In the open-channel conformation, the position of the S4–S5 linker forces the S6 helix to form a kink near the cytosolic surface (blue in Figure 22-14a) and the pore inside, near the cytosolic surface, is open. The pore’s 1.2-nm diameter is sufficient to accommodate hydrated K+ ions (see Figure 22-14c).
When the cell membrane is repolarized and the voltage sensor moves toward the cytosolic membrane surface, the S4–S5 linkers (orange in Figure 22-14b) are twisted down, toward the inside of the cell. The S6 helices are consequently straightened, squeezing the bottom of the channel closed. Thus the gate is composed of the cytosol-facing ends of the S5 and S6 helices, where the pore is narrowest.
Although voltage-gated K+ and voltage-gated Na+ channels share similar voltage sensor and pore structures, the structure of their ion selectivity filters and the way that they conduct ions differ significantly (as also discussed in Chapter 11). The selectivity filter of the Na+ channel is much larger than that of the K+ channel, even though the diameter of a Na+ ion (0.102 nm) is smaller than that of a K+ ion (0.138 nm). The pore of K+ channels contains conserved amino acids that form a lining of carbonyl oxygen atoms (see Figure 11-21). As K+ ions enter the pore, these oxygen atoms replace its waters of hydration; the smaller Na+ ions would be too small to interact with the backbone carbonyls of the K+ channel pore. In contrast, Na+ ions pass through the Na+ channel pore as water-hydrated ions. The pore of the Na+ channel is lined by conserved negatively charged amino acids and is large enough to fit a single, water-hydrated Na+ ion, with the positively charged Na+ ion interacting with the negatively charged pore residues through its inner shell of bound water molecules. The hydrated K+ ion is too large to fit through this pore.
The topical anesthesia lidocaine, which is commonly used to reduce pain during dental procedures or during minor surgeries (e.g., to suture cuts), works by blocking the flow of Na+ ions through the voltage-gated Na+ channel. Lidocaine binds to amino acid residues that line the channel pore and prevent the influx of Na+, and thus the generation of an action potential. The binding sites for lidocaine are only available in the open state of the channel, and binding of lidocaine appears to lock the channel in the open but occluded state.