Viral DNA Is Integrated into the Host-Cell Genome in Some Nonlytic Viral Growth Cycles
Some bacterial viruses, called temperate phages, can establish a nonlytic association with their host cell that does not kill the cell. For example, when bacteriophage λ infects E. coli, most of the time it causes a lytic infection. Occasionally, however, the viral DNA is integrated into the host-cell chromosome rather than being replicated. The integrated viral DNA, called a prophage, is replicated as part of the host cell’s DNA from one host-cell generation to the next. This phenomenon is referred to as lysogeny. If the host cell suffers extensive damage to its DNA from UV light, the prophage DNA is activated, leading to its excision from the host-cell chromosome, entrance into the lytic cycle, and subsequent production and release of progeny virions before the host cell dies.
The genomes of a number of animal viruses can also integrate into the host-cell genome. Among the most important of these are the retroviruses, which are enveloped viruses with a genome consisting of two identical strands of RNA. These viruses are so named because their RNA genome acts as a template for the formation of a DNA molecule—a flow of genetic information that is opposite to the more common transcription of DNA into RNA. In the retroviral life cycle (Figure 5-48), a viral enzyme called reverse transcriptase initially copies the viral RNA genome into single-stranded DNA that is complementary to the viral RNA; the same enzyme then catalyzes the synthesis of a complementary DNA strand. (This complex reaction is detailed in Chapter 8, where we consider closely related intracellular parasites called retrotransposons.) The resulting double-stranded DNA is integrated into the chromosomal DNA of the infected cell by an integrase enzyme in the virion. Finally, the integrated DNA, called a provirus, is transcribed by the host cell’s own RNA polymerase into RNA, which is either translated into viral proteins or packaged within virion capsid proteins to form progeny virions that are released by budding from the host-cell membrane. Because most retroviruses do not kill their host cells, infected cells can replicate, producing daughter cells with integrated proviral DNA. These daughter cells continue to transcribe the proviral DNA and bud progeny virions.
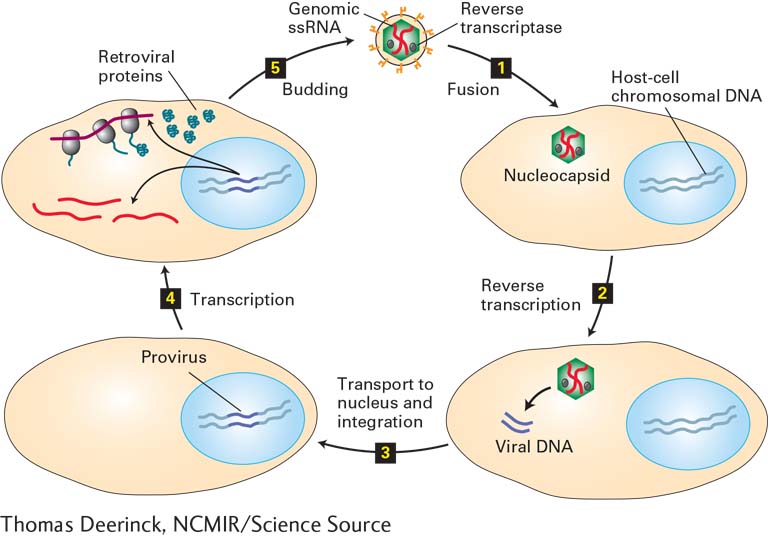
FIGURE 5-48 Retroviral life cycle. Retroviruses have a genome of two identical copies of single-stranded RNA and an outer envelope. Step 1: After viral glycoproteins in the retroviral envelope interact with a specific host-cell membrane protein, the envelope fuses directly with the plasma membrane, allowing entry of the nucleocapsid into the cytoplasm of the cell. Step 2: Viral reverse transcriptase and other proteins copy the viral ssRNA genome into a double-stranded DNA. Step 3: The viral dsDNA is transported into the nucleus and integrated into one of many possible sites in the host-cell chromosomal DNA. For simplicity, only one host-cell chromosome is depicted. Step 4: The integrated viral DNA (provirus) is transcribed by the host-cell RNA polymerase, generating viral mRNAs (dark red) and viral genomic RNA molecules (bright red). The host-cell machinery translates the viral mRNAs into glycoproteins and nucleocapsid proteins. Step 5: Progeny virions then assemble and are released by budding, as illustrated in Figure 5-46.
Some retroviruses contain cancer-causing genes (oncogenes), and cells infected by such retroviruses are oncogenically transformed into tumor cells. Studies of oncogenic retroviruses (mostly viruses of birds and mice) have revealed a great deal about the processes that lead to transformation of a normal cell into a cancer cell (see Chapter 24).
Among the known human retroviruses are human T-cell lymphotrophic virus (HTLV), which causes a form of leukemia, and human immunodeficiency virus (HIV-1), which causes acquired immune deficiency syndrome (AIDS). Both of these viruses can infect only specific cell types, primarily certain cells of the immune system and, in the case of HIV-1, some central nervous system neurons and glial cells. Only those cells have cell-surface receptors that interact with viral envelope proteins. Unlike most other retroviruses, HIV-1 eventually kills its host cells. The eventual death of large numbers of immune-system cells results in the defective immune response characteristic of AIDS.
Some DNA viruses can also integrate into a host-cell chromosome. One example is the human papillomaviruses (HPVs), which most commonly cause warts and other benign skin lesions. These viruses replicate without integrating into a host-cell chromosome. However, the genomes of certain HPV serotypes occasionally integrate into the chromosomal DNA of infected cervical epithelial cells. Unlike retroviruses and temperate bacteriophages, this integration is not carried out by viral proteins, but rather is the consequence of host-cell DNA repair processes. Integration is a dead-end for HPV. The integrated viral DNA cannot replicate and generate progeny virions. But oncogenic (cancer-causing) viral proteins can be expressed from the integrated viral genome, initiating development of cervical cancer. Routine Pap smears can detect cells in the early stages of the transformation process initiated by HPV integration, permitting effective treatment before cancer develops. A vaccine for the types of HPV associated with cervical cancer has been developed and can protect against the initial infection by these viruses, and consequently, against development of cervical cancer. However, once an individual is infected with these HPVs, the “window of opportunity” is missed, and the vaccine does not protect against the development of cancer. Because the vaccine is not 100 percent effective, even vaccinated women should have regular Pap smears.