Mobile DNA Elements Have Significantly Influenced Evolution
Although mobile DNA elements appear to have no direct function other than to maintain their own existence, their presence has had a profound effect on the evolution of modern-day organisms. As mentioned earlier, about half the spontaneous mutations in Drosophila result from insertion of a mobile DNA element into or near a transcription unit. In mammals, mobile elements cause a much smaller proportion of spontaneous mutations: about 10 percent in mice, and only 0.1–0.2 percent in humans. Still, mobile elements have been found in mutant alleles associated with several human genetic diseases. For example, insertions into the clotting factor IX gene cause hemophilia, and insertions into the gene encoding the muscle protein dystrophin lead to Duchenne muscular dystrophy. The genes encoding clotting factor IX and dystrophin are both on the X chromosome. Consequently, disease resulting from a transposition into these genes occurs primarily in males, in which there is no second copy of the normal gene to complement the resulting mutation.
In lineages leading to higher eukaryotes, homologous recombination between mobile DNA elements dispersed throughout ancestral genomes may have generated gene duplications and other DNA rearrangements during evolution (see Figure 8-2b). For instance, cloning and sequencing of the β-globin gene clusters from various primate species has provided strong evidence that the human HGB1 and HGB2 genes encoding fetal β-globins arose from an unequal homologous crossover between two L1 elements flanking an ancestral globin gene. Subsequent divergence of such duplicated genes could lead to the acquisition of distinct, beneficial functions by each member of a gene family. Unequal crossing over between mobile elements located within introns of a particular gene could lead to the duplication of exons within that gene (see Figure 8-2a). This process most likely influenced the evolution of genes that contain multiple copies of similar exons encoding similar protein domains, such as the fibronectin gene (see Figure 5-16).
Some evidence suggests that during the evolution of higher eukaryotes, recombination between mobile DNA elements (e.g., Alu elements) in introns of two separate genes also occurred, generating new genes made from novel combinations of preexisting exons (Figure 8-18). This evolutionary process, termed exon shuffling, may have occurred during the evolution of the genes encoding tissue plasminogen activator, the Neu receptor, and epidermal growth factor, all of which contain an EGF domain (see Figure 3-12). In this case, exon shuffling presumably resulted in the insertion of an EGF domain–encoding exon into an intron of the ancestral form of each of these genes.
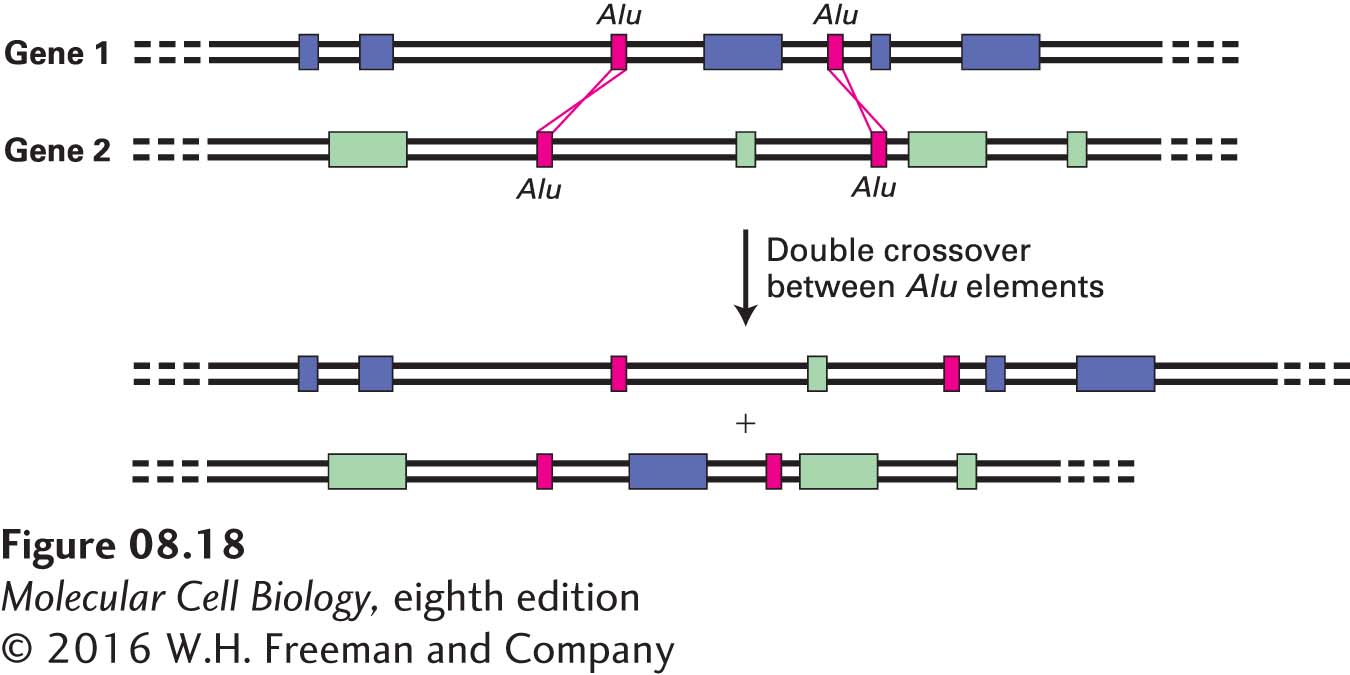
FIGURE 8-18 Exon shuffling via recombination between homologous interspersed repeats. Recombination between interspersed repeats in the introns of separate genes produces transcription units with new combinations of exons. In the example shown here, a double crossover between two sets of Alu repeats results in an exchange of exons between two genes.
Both DNA transposons and LINE retrotransposons have been shown to occasionally carry unrelated flanking sequences—including exons—when they transpose to new sites by the mechanisms diagrammed in Figure 8-19. These mechanisms probably also contributed to exon shuffling during the evolution of contemporary genes.
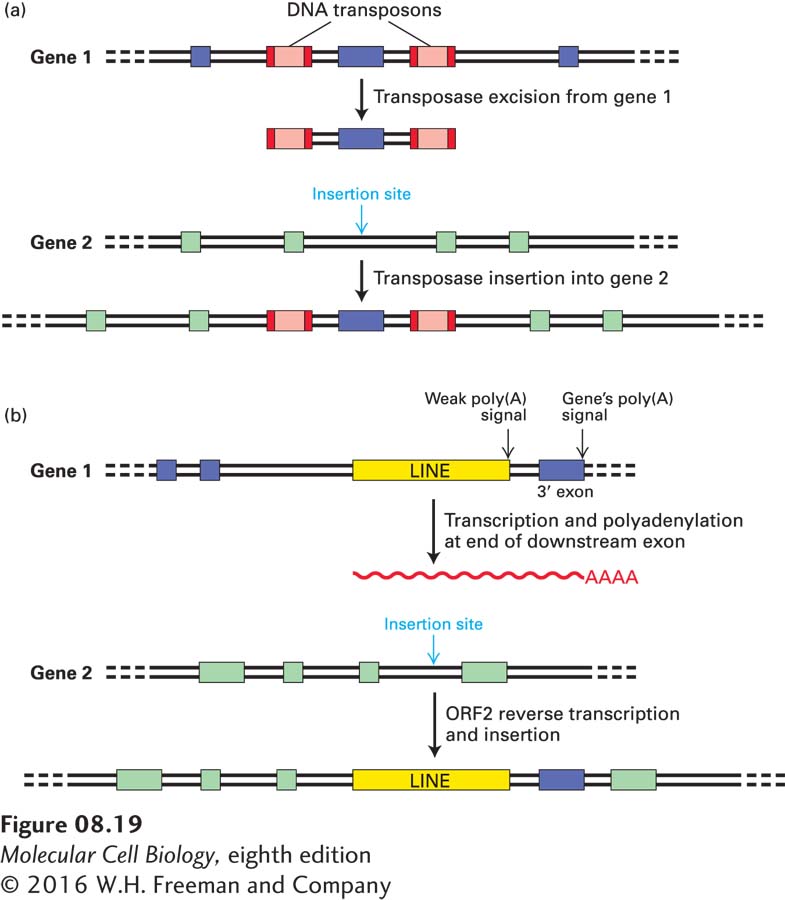
FIGURE 8-19 Exon shuffling by transposition. (a) Transposition of an exon flanked by homologous DNA transposons into an intron on a second gene. As we saw in Figure 8-10, step 1, transposase can recognize and cleave the DNA at the ends of the transposon inverted repeats. In gene 1, if the transposase cleaves at the left end of the transposon on the left and at the right end of the transposon on the right, it can transpose all the intervening DNA, including the exon from gene 1, to a new site in an intron of gene 2. The net result is an insertion of the exon from gene 1 into gene 2. (b) Integration of an exon into another gene via LINE transposition. Some LINEs have weak poly(A) signals. If such a LINE is in the 3′-most intron of gene 1, during transposition its transcription may continue beyond its own poly(A) signals and extend into the 3′ exon, transcribing the cleavage and polyadenylation signals of gene 1 itself. This RNA can then be reverse-transcribed and integrated by the LINE ORF2 protein (see Figure 8-17) into an intron on gene 2, introducing a new 3′ exon (from gene 1) into gene 2.
In addition to causing changes in coding sequences, recombination between mobile elements and transposition of DNA adjacent to mobile elements probably played a significant role in the evolution of regulatory sequences that control gene expression. As noted earlier, eukaryotic genes have transcription-control regions called enhancers that can operate over distances of tens of thousands of base pairs. The transcription of many genes is controlled through the combined effects of several enhancer elements. Insertion of mobile elements near such transcription-control regions probably contributed to the evolution of new combinations of enhancer sequences. These combinations, in turn, control which specific genes are expressed in particular cell types and the amount of the encoded protein produced in modern organisms, as we discuss in the next chapter.
These considerations suggest that the early view of mobile DNA elements as completely selfish molecular parasites misses the mark. Rather, these elements have contributed profoundly to the evolution of higher organisms by promoting (1) the generation of gene families via gene duplication, (2) the creation of new genes via shuffling of preexisting exons, and (3) the formation of more complex regulatory regions that provide multifaceted control of gene expression. Today researchers are attempting to harness transposition mechanisms to insert therapeutic genes into patients as a form of gene therapy.
A process analogous to that shown in Figure 8-19a is largely responsible for the rapid spread of antibiotic resistance among pathogenic bacteria, a major problem in modern medicine. Bacterial genes encoding enzymes that inactivate antibiotics (drug resistance genes) have become flanked by insertion sequences, generating drug resistance transposons. The widespread use of antibiotics in medicine, often unnecessarily in the treatment of viral infections, on which they have no effect, and to prevent infections in healthy agricultural animals has led to positive selection on such drug resistance transposons, which have inserted into conjugating plasmids. Conjugating plasmids encode proteins that result in their replication and transfer to other bacterial cells—even cells of other related bacterial species—through a complex macromolecular tube called a pilus. These plasmids, called R factors (for drug resistance), can contain multiple drug resistance genes introduced by transposition and selected in environments, such as hospitals, where antibiotics are used to sterilize surfaces. These R factors have led to the rapid spread of resistance to multiple antibiotics between pathogenic bacteria. Coping with the spread of R factors is a major challenge for modern medicine.