Three Functional Elements Are Required for Replication and Stable Inheritance of Chromosomes
Although eukaryotic chromosomes differ in length and number among species, cytogenetic studies have shown that they all behave similarly at the time of cell division. Moreover, any eukaryotic chromosome must contain three functional elements in order to replicate and segregate correctly: (1) replication origins at which DNA polymerases and other proteins initiate synthesis of DNA (see Figures 5-31 and 5-33); (2) the centromere, the constricted region required for proper segregation of daughter chromosomes; and (3) the two ends, or telomeres. The yeast transformation studies depicted in Figure 8-41 demonstrated the functions of these three chromosomal elements and established their importance for chromosome function.
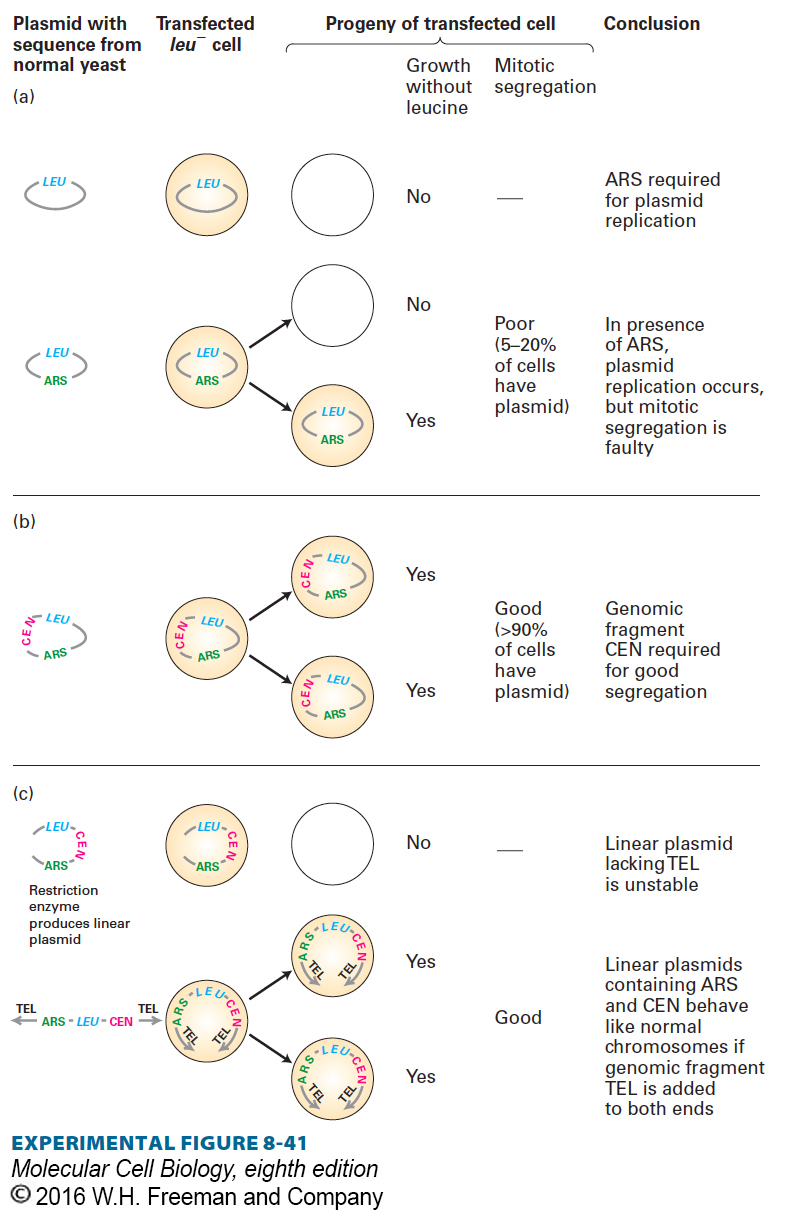
EXPERIMENTAL FIGURE 8-41 Yeast transformation experiments were used to identify the functional chromosomal elements necessary for normal chromosome replication and segregation. In these experiments, plasmids containing the LEU gene from normal yeast cells are constructed and introduced into leu− cells by transfection. If the plasmid is maintained in the leu− cells, they are transformed to LEU+ cells by the LEU gene on the plasmid and can form colonies on medium lacking leucine. (a) Sequences that allow autonomous replication (ARS) of a plasmid were identified because their insertion into a plasmid vector containing a cloned LEU gene resulted in a high frequency of transformation to LEU+. However, even plasmids with ARS exhibit poor segregation during mitosis and therefore do not appear in each of the daughter cells. (b) When randomly broken pieces of yeast DNA are inserted into plasmids containing ARS and LEU, some of the subsequently transfected cells produce large colonies, indicating that a high rate of mitotic segregation among their plasmids is facilitating the continuous growth of daughter cells. The DNA recovered from plasmids in these large colonies contains yeast centromere (CEN) sequences. (c) When leu− yeast cells are transfected with linearized plasmids containing LEU, ARS, and CEN, no colonies grow. Addition of telomere (TEL) sequences to the ends of the linear DNA gives the linearized plasmids the ability to replicate as new chromosomes that behave very much like a normal chromosome in both mitosis and meiosis. See A. W. Murray and J. W. Szostak, 1983, Nature 305:89, and L. Clarke and J. Carbon, 1985, Ann. Rev. Genet. 19:29.
As discussed in Chapter 5, replication of DNA begins from sites that are scattered throughout eukaryotic chromosomes. The yeast genome contains many 100-bp sequences, called autonomously replicating sequences (ARSs), that act as replication origins. The observation that insertion of an ARS into a circular plasmid allows the plasmid to replicate in yeast cells provided the first functional identification of replication origins in eukaryotic DNA (Figure 8-41a).
Even though circular ARS-containing plasmids can replicate in yeast cells, only about 5–20 percent of progeny cells contain the plasmid because mitotic segregation of the plasmids is faulty. However, plasmids that also carry a CEN sequence, derived from the centromeres of yeast chromosomes, segregate equally, or nearly so, to both mother and daughter cells during mitosis (Figure 8-41b).
If circular plasmids containing an ARS and a CEN sequence are cut once with a restriction enzyme, the resulting linear plasmids do not transform yeast cells generating LEU+ colonies that grow on medium lacking leucine unless they contain special telomeric (TEL) sequences ligated to their ends (Figure 8-41c). The first successful experiments involving transfection of yeast cells with linear plasmids were achieved by using the ends of a DNA molecule that was known to replicate as a linear molecule in the ciliated protozoan Tetrahymena. During part of the life cycle of Tetrahymena, much of the nuclear DNA is repeatedly copied in short pieces to form a so-called macronucleus. One of these repeated fragments was identified as a dimer of ribosomal DNA, the ends of which contained a repeated sequence (G4T2)n. When a section of this repeated TEL sequence was ligated to the ends of linear yeast plasmids containing ARS and CEN, replication and good segregation of the linear plasmids occurred. This first cloning and characterization of telomeres garnered the Nobel Prize in Physiology or Medicine in 2009.