DNA Methylation Represses Transcription
As mentioned earlier, most promoters in mammals fall into the CpG island class. Active CpG island promoters have Cs in their CG sequences that are unmethylated. Unmethylated CpG island promoters have reduced affinity for histone octamers, but nucleosomes immediately neighboring the unmethylated promoters are modified by histone H3 lysine 4 di- or trimethylation and are associated with Pol II molecules that are paused during transcription of both the sense and antisense template DNA strands, as discussed earlier (see Figures 9-18 and 9-19). Recent research indicates that methylation of histone H3 lysine 4 occurs in mouse cells because a protein named Cfp1 (CXXC finger protein 1) binds unmethylated CpG-rich DNA through a zinc-finger domain (CXXC) and associates with a histone methylase specific for histone H3 lysine 4 (Setd1). Chromatin-remodeling complexes and the general transcription factor TFIID, which initiates Pol II preinitiation complex assembly (see Figure 9-19), associate with nucleosomes bearing the H3 lysine 4 trimethyl mark, promoting Pol II transcription initiation.
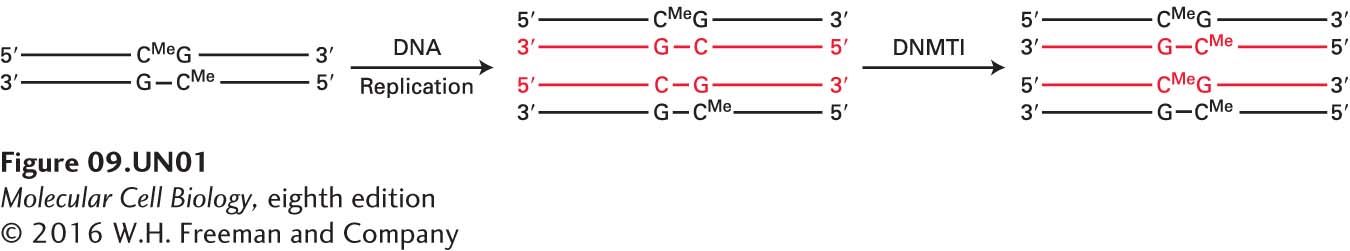
In differentiated cells, however, a small percentage of specific CpG island promoters, depending on the cell type, have CpGs marked by 5-methyl C. This modification of CpG island DNA triggers chromatin condensation. A family of proteins that bind to DNA that is rich in 5-methyl C–modified CpGs (called methyl CpG-binding proteins, or MBDs) bind to the marked promoters and associate with histone deacetylases and repressive chromatin-remodeling complexes that condense chromatin, resulting in transcriptional repression. The 5-methyl C is added to the CpGs by DNA methyl transferases named DNMT3a and DNMT3b. They are referred to as de novo DNA methyl transferases because they methylate an unmethylated C. Much remains to be learned about how DNMT3a and b are directed to specific CpG islands. But once they have methylated a DNA sequence, methylation at that C is passed on through DNA replication through the action of the ubiquitous maintenance methyl transferase DNMT1:
(red indicates daughter strands). As a consequence, once a CpG island promoter is methylated by DNMT3a or b, it continues to be methylated by DNMT1 in subsequent daughter cells. Consequently, the promoter remains repressed in all subsequent daughter cells through interactions with MBDs, even after the stimulus for the initial C-methylation by DNMT3a or b has ceased. Therefore, repression of C-methylated promoters is inherited through cell division. This mechanism of epigenetic repression is being intensely investigated because tumor-suppressor genes encoding proteins that function to suppress the development of cancer are often inactivated in cancer cells by abnormal CpG methylation of their promoter regions, as discussed further in Chapter 24.