Microtubules and microfilaments are dynamic structures.
When we think of the parts of a skeleton, we are inclined to think of our bones. Unlike our bones, which undergo remodeling but do not change rapidly, microtubules and microfilaments are dynamic. They become longer by the addition of subunits to their ends, and become shorter by the loss of subunits.
The rate at which protein subunits are added depends on the concentrations of tubulin and actin in that region of the cell. At high concentrations of subunits, microtubules and microfilaments can become longer at both ends, although the subunits are assembled more quickly on one end than the other. The faster-
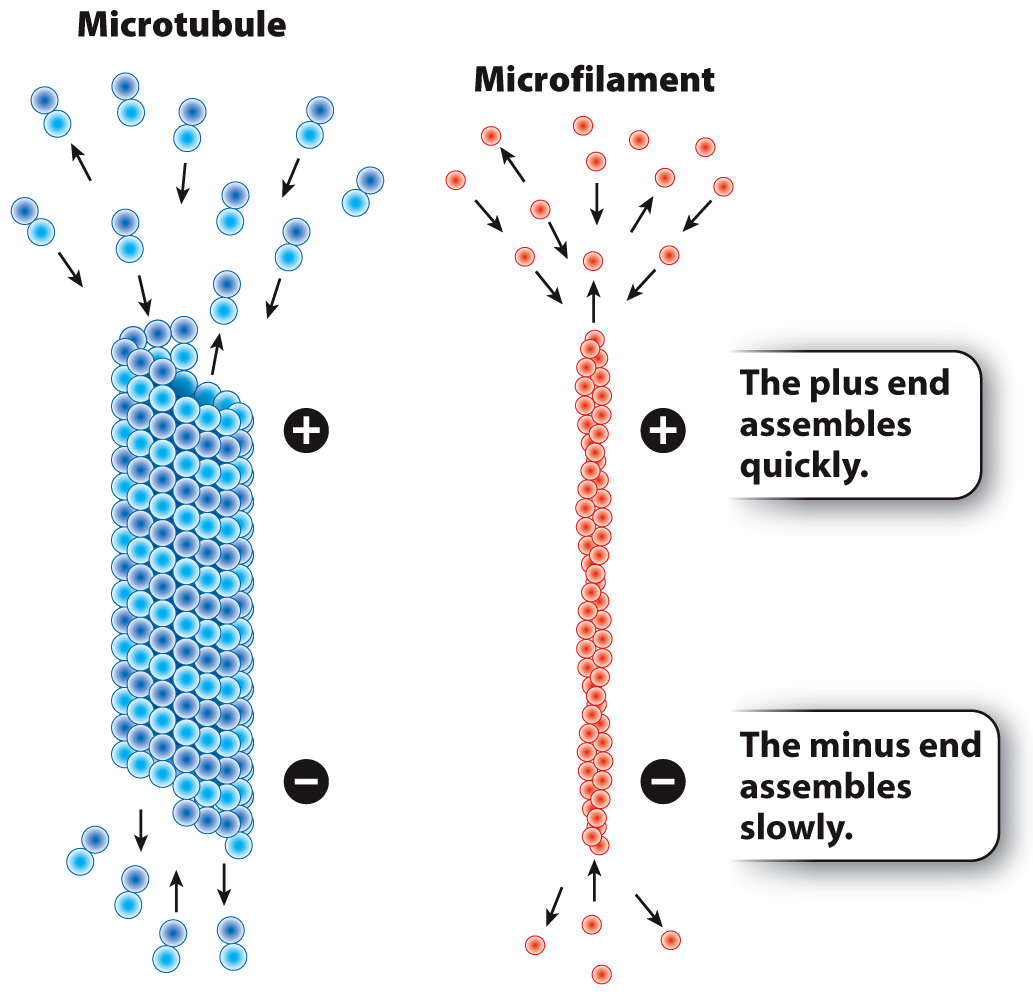
The dynamic nature of microfilaments and microtubules is important for some of their functions. For example, some forms of cell movement, such as a single-
202
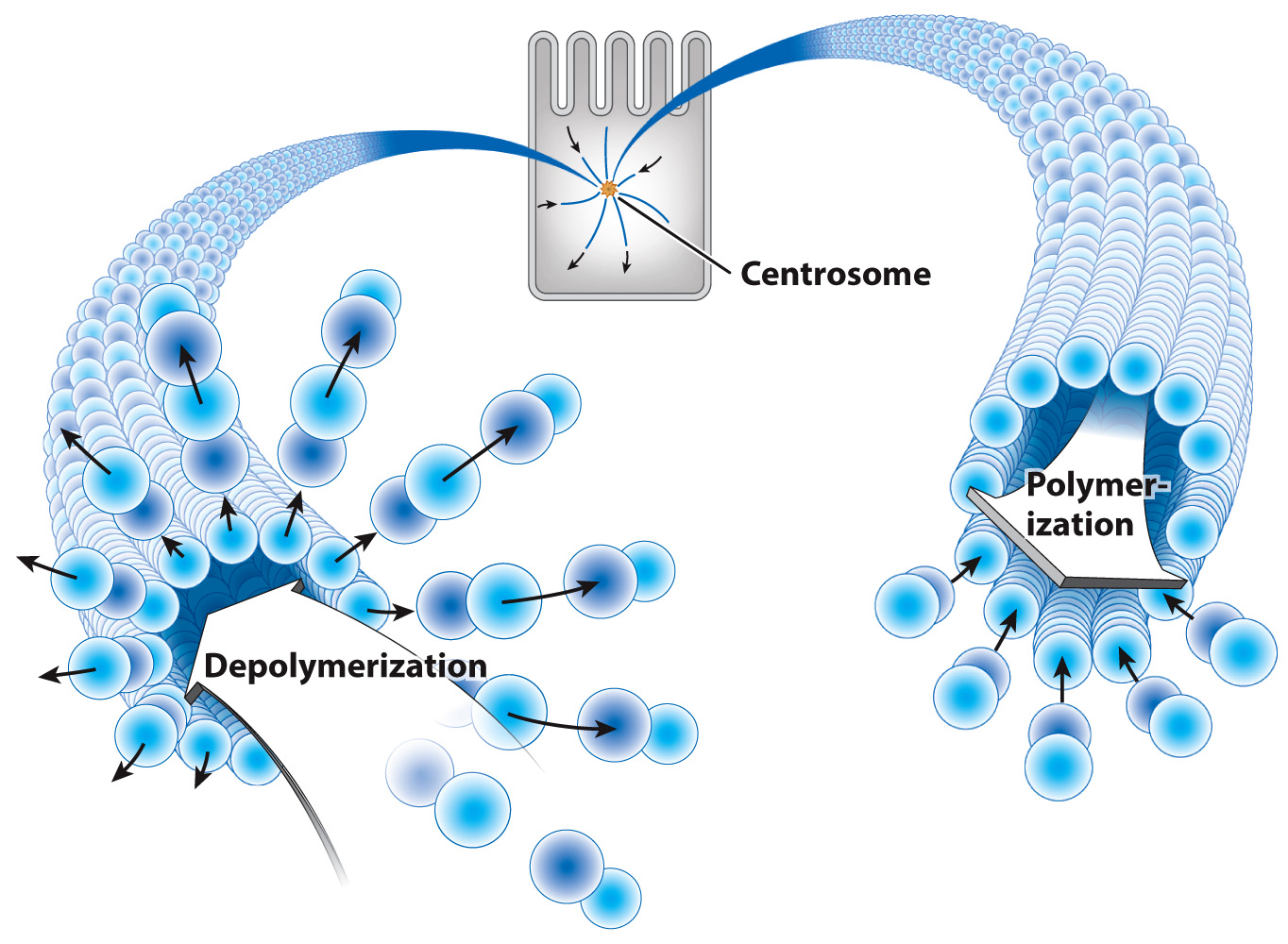
Microtubules make up the spindles that attach to chromosomes during cell division. In this case, the ability of spindle microtubules to “explore” the space of the cell and encounter chromosomes is driven by a unique property of microtubules: Their plus ends undergo seemingly random cycles of rapid depolymerization followed by slower polymerization. These cycles of depolymerization and polymerization are called dynamic instability (Fig. 10.6). They allow spindle microtubules to quickly find and attach to chromosomes during cell division.