A few genetic differences are beneficial.
While many mutations are neutral, or nearly so, and many others are harmful, some mutations are beneficial. In human populations, beneficial mutations are often discovered through their effects in protecting against infectious disease. The most widely known example is probably the sickle-
By means of its surface glycoprotein (a product of the env region in the annotated HIV genome shown in Fig. 13.7), HIV combines with a cell-
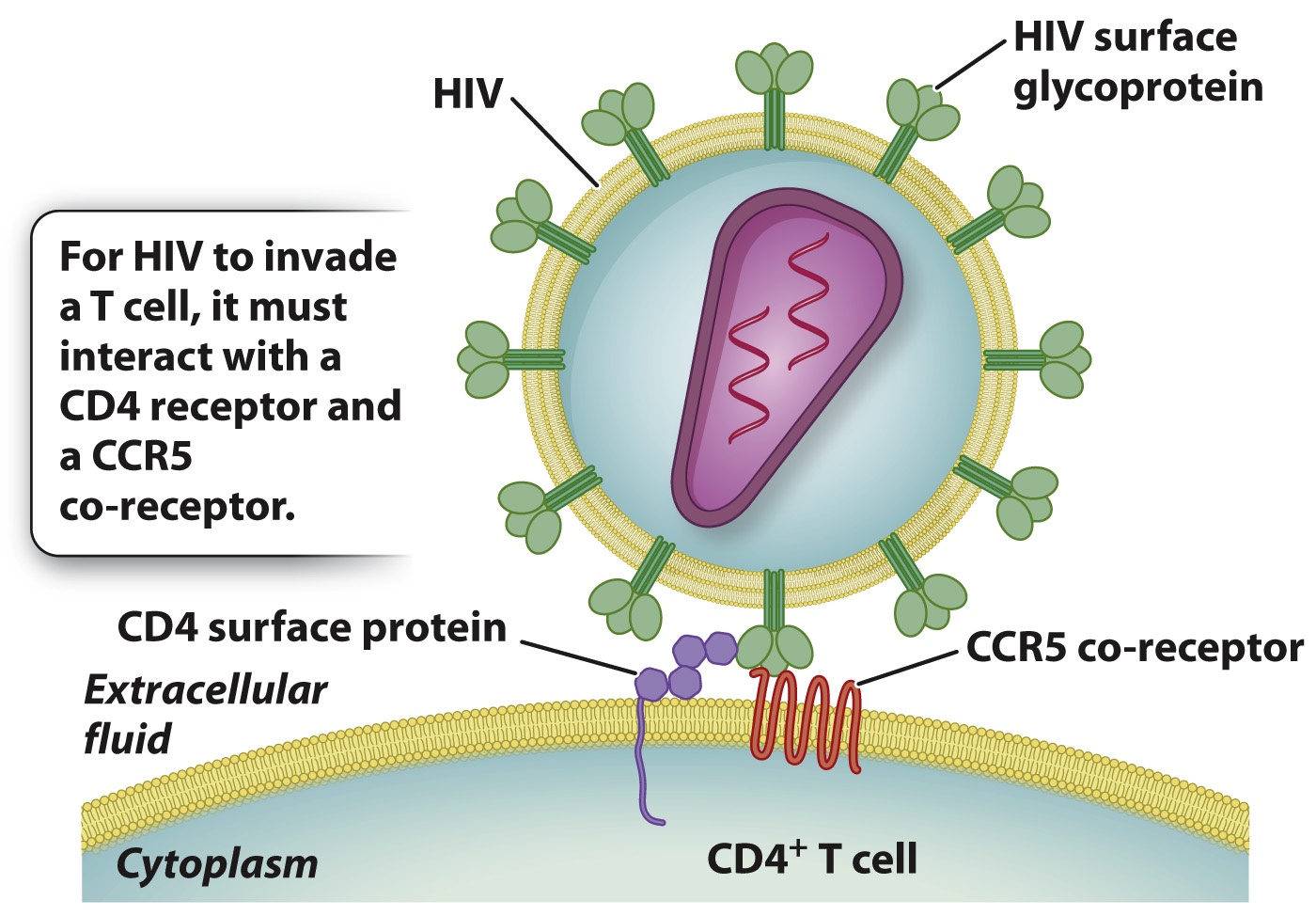
A beneficial effect of a particular mutation in the CCR5 gene was discovered in studies focusing on HIV patients whose infection had not progressed to full-

314
The effect of the Δ32 allele is pronounced. In individuals with the homozygous Δ32/Δ32 genotype, HIV progression to AIDS is rarely observed. There is some protection even in individuals with heterozygous Δ32 genotypes, where progression to AIDS is delayed by an average of about 2 years after infection by HIV.
Much has been written about the evolutionary history of the Δ32 allele. It is found almost exclusively in European populations, where the frequency of heterozygous genotypes ranges from 10% to 25%. The narrow geographical distribution was originally interpreted to mean that the allele was selected over time because it provided protection against some other infectious agent that also interacted with the CCR5 protein.
Beneficial mutations not only provide protection against disease. Many beneficial mutations permit organisms to become better adapted to their environment. For example, certain birds, such as Rüppell’s vulture, commonly fly at altitudes of 20,000 feet and sometimes much higher. This feat is possible because of mutations in the structure of hemoglobin that allow hemoglobin to bind oxygen with high affinity, even at the low pressure of oxygen high in the atmosphere. These mutations were selected and passed on generation after generation, making the birds well adapted to flying at high altitudes.