In crosses, one of the traits was dominant in the offspring.
Mendel began his studies with crosses between true-
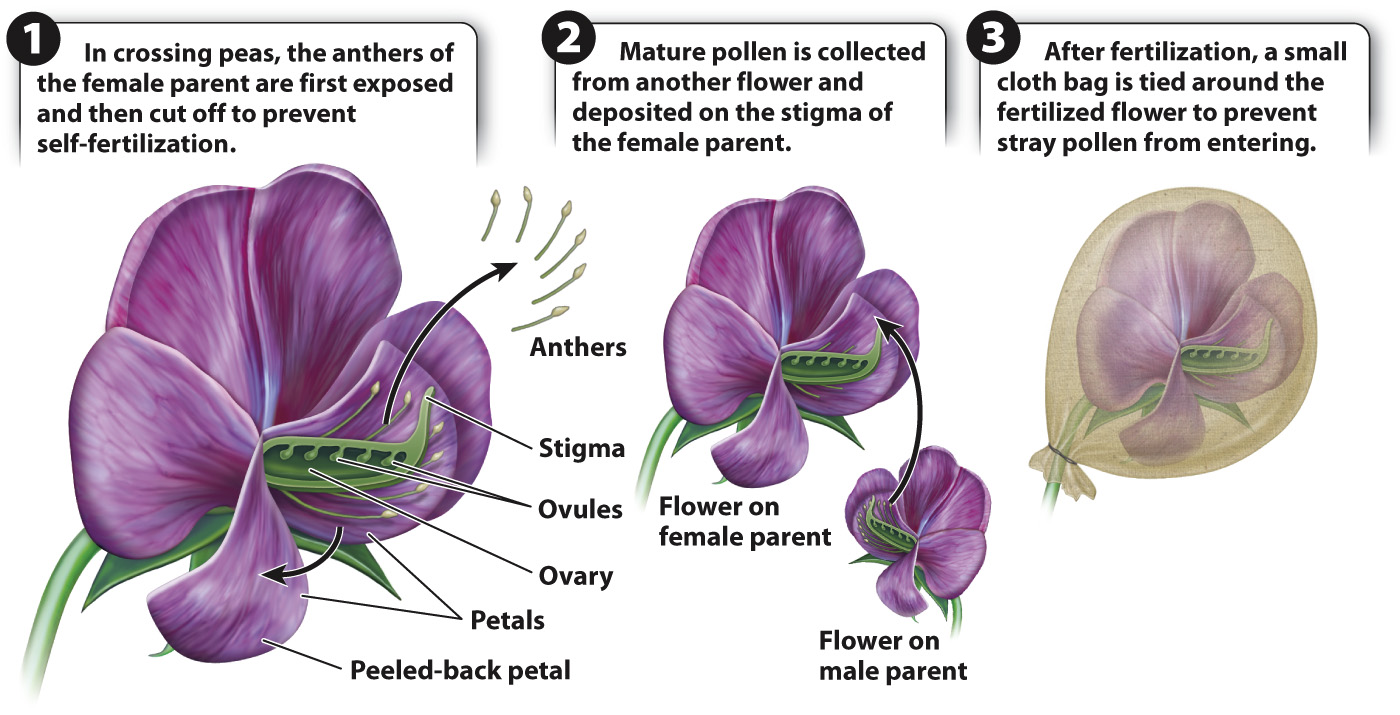
329
For each of the seven pairs of contrasting traits, true-

Crosses like these, in which the expressions of the trait in the female and male parents are interchanged, are known as reciprocal crosses. Mendel showed that reciprocal crosses yielded the same result for each of his seven pairs of contrasting traits.
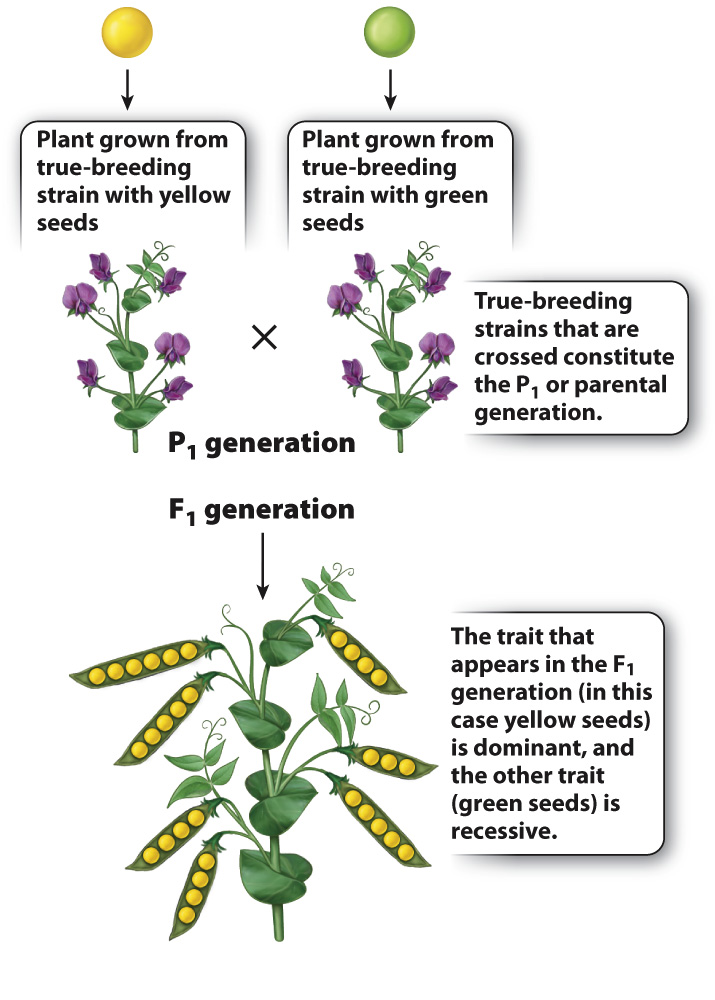
Moreover, for each pair of contrasting traits, only one of the traits appeared in the F1 generation. For simple crosses involving two parents that are true breeding for different traits, the trait that appears in the F1 generation is said to be dominant, and the contrasting trait that does not appear is said to be recessive. For each pair of traits illustrated in Fig. 16.3, the dominant trait is shown on the left, and the recessive on the right. Thus, yellow seed is dominant to green seed, round seed is dominant to wrinkled seed, and so forth.
Mendel explained these findings by supposing that there is a hereditary factor for yellow seeds and a different hereditary factor for green seeds, likewise a hereditary factor for round seeds and a different one for wrinkled seeds, and so on. We now know that the hereditary factors that result in contrasting traits are different forms of a gene that affect the trait. The different forms of a gene are called alleles. The alleles of a gene are variations of the DNA sequence of a gene that occupies a particular region along a chromosome. The particular combination of alleles present in an individual constitutes its genotype, and the expression of the trait in the individual constitutes its phenotype. As noted, Mendel found that, for each of his traits, one phenotype was dominant and the other phenotype was recessive.
330
In the cross between true-
The molecular basis of dominance in this case is the fact that, in diploid organisms, only one copy of a gene is needed to carry out its normal function. The yellow color in pea seeds results from an enzyme that breaks down green chlorophyll, allowing yellow pigments to show through. Green seeds result from a mutation in this gene that inactivates the enzyme, and so the green chlorophyll is retained. In an F1 hybrid, such as that in Fig. 16.5, which receives a nonmutant gene from one parent and a mutant gene from the other, the seeds are yellow because one copy of the nonmutant gene produces enough of the enzyme to break down the chlorophyll to yield a yellow seed.