Complex traits are affected by multiple genes.
Most complex traits are affected by many genes, in contrast to Mendel’s traits, which are primarily affected by only one. Therefore, in complex traits the familiar phenotypic ratios such as 3:1 that Mendel saw when he crossed two true-breeding strains with contrasting characters are not observed. For complex traits, the effects of individual genes are obscured by variation in phenotype that is due to multiple genes affecting the trait and also due to the environment. The number of genes affecting complex traits is usually so large that different genotypes can have very similar phenotypes, which also makes it difficult to see the effects of individual genes on a trait.
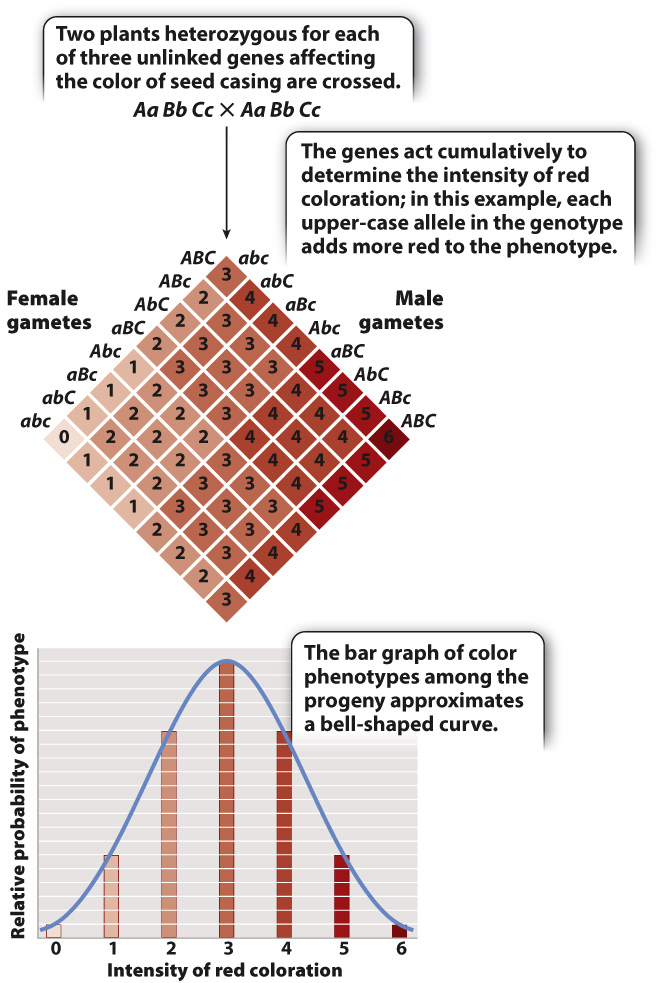
FIG. 18.4 Multiple genes contributing to a complex trait. Three unlinked genes, each with two alleles, influence the intensity of red coloration of the seed casing in wheat.
In a few traits, however, the effects of the environment are minor and the number of genes is small, and in these cases the genetic basis of the trait can be analyzed. A classic example, studied by Herman Nilsson-Ehle about a century ago, concerns the color of seed casing in wheat (which give the seeds their color), which ranges from nearly white to dark red (Fig. 18.4). His experiment demonstrated that complex traits are subject to the same laws that Mendel worked out for single-gene traits, but that the inheritance patterns are more difficult to see because of the number of genes involved.
In studying seed color in true-breeding varieties and their first-generation and second-generation hybrids, Nilsson-Ehle realized that the relative frequencies of different shades of red color could be explained by the effects of three genes that undergo independent assortment, as illustrated by the Punnett square shown in Fig. 18.4. Each of the genes has two alleles, designated by combinations of upper-case and lower-case letters. In the development of seed color, each upper-case allele in a genotype intensifies the red coloration in an additive fashion. Altogether there are seven possible phenotypes, ranging from a phenotype of 0 (nearly colorless, genotype aa bb cc) to 6 (dark red, genotype AA BB CC).
Nilsson-Ehle first crossed true-breeding dark red plants (AA BB CC) with true-breeding colorless plants (aa bb cc) to obtain the hybrids with genotype Aa Bb Cc. He then crossed the hybrids together, which is the cross shown at the top of Fig. 18.4. Because the three genes show independent assortment (Chapter 16), the distribution of phenotypes expected in progeny from the cross Aa Bb Cc × Aa Bb Cc is as shown in the Punnett square. The bar graph below the Punnett square shows the seed-color phenotypes and their relative proportions, from which Nilsson-Ehle inferred three genes with independent assortment. The distribution of seed-color phenotypes is approximated by a bell-shaped curve known as a normal distribution. The phenotypes of many complex traits, including human height, conform to the normal distribution. Nilsson-Ehle’s seed-color case is exceptional in that virtually all complex traits are affected by many more genes than three (in the case of human height, hundreds, as discussed below).
When differences in phenotype due to the environment can be ignored, the genetic variation affecting complex traits can be detected more easily. And when studying inbred lines, differences in phenotype due to genotype can be ignored because all individuals have the same genotype, and the effects of environment can be observed. In most cases, however, both genetic variation and environmental variation among individuals are present, and it is difficult to quantify how much variation in phenotype is due to genes and how much is due to environment.
It’s important to point out that complex traits are not really more “complex” than any other biological trait. The term is used merely to imply that both genetic factors and environmental factors contribute to variation in phenotype among individuals. Just as there are environmental factors that affect complex traits such as height, so there are genetic factors that affect height. Similarly, just as an environmental risk factor increases the likelihood of a common disease, so does a genetic risk factor predispose an individual to the condition. For example, the human gene ApoE encodes a protein that helps transport fat and cholesterol. Certain alleles of ApoE are associated with high levels of cholesterol, and one particular allele is a genetic risk factor for Alzheimer’s disease, the most common serious form of age-related loss of cognitive ability. Lifestyle environmental factors such as diet, physical exercise, and mental stimulation are also thought to play a role in the risk of Alzheimer’s disease.