The lactose operon is also positively regulated by CRP–cAMP.
Fig. 19.16 and Fig. 19.17 show how the ability of the repressor to bind with either the operator (in the absence of lactose) or with the inducer (in the presence of lactose) provides a simple and elegant way for the bacterial cell to transcribe the genes needed for lactose utilization only in the presence of lactose. The elucidation of these interactions was as far as the research tools used by Jacob and Monod could take them. Since the original experiments, the lactose operon has been studied in much greater detail and additional levels of regulation have been discovered.
One of these additional levels involves the CRP binding site shown in Fig. 19.15, which in Fig. 19.19 is occupied by a protein called the CRP–cAMP complex. The CRP–cAMP complex is a positive regulator of the lactose operon, which you will recall is a protein that activates gene expression upon binding DNA. “CRP” stands for “cAMP receptor protein.” The role of CRP–cAMP is to provide another level of control of transcription that is more sensitive to the nutritional needs of the cell than the level of control provided by the presence or absence of lactose. E. coli can utilize many kinds of molecules as sources of energy. When more than one type of energy source is available in the environment, certain sources are used before others. For example, glucose is preferred to lactose, and lactose is preferred to glycerol. The CRP–cAMP complex helps regulate which compounds are utilized.
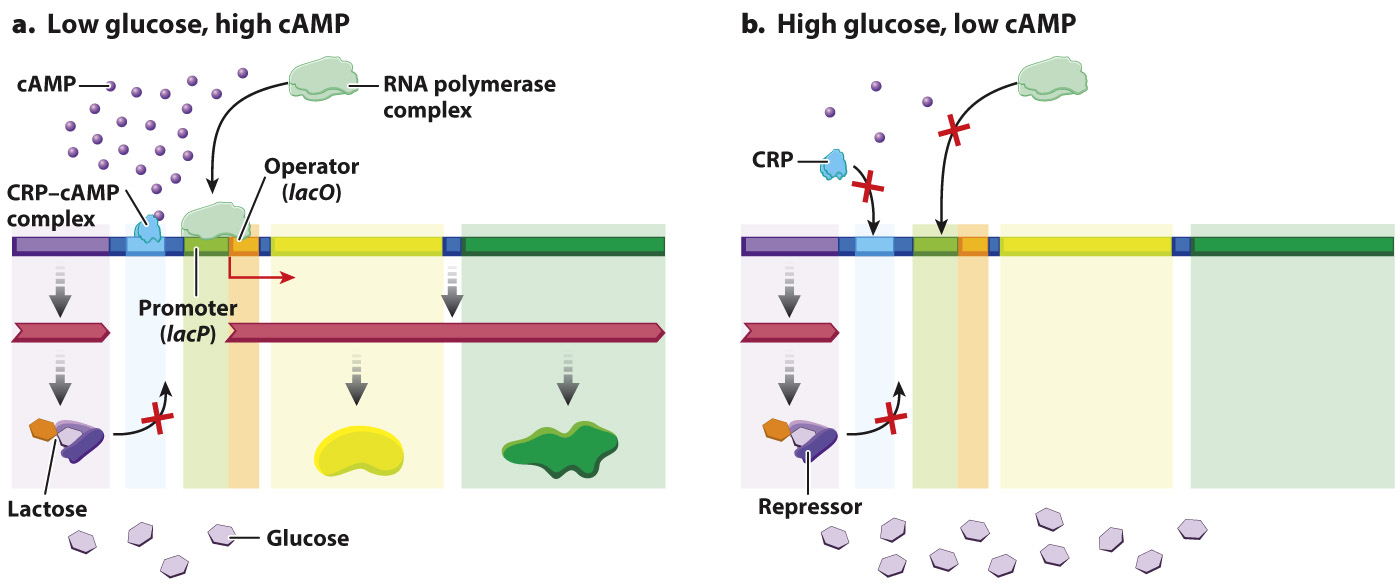
FIG. 19.19 The CRP–cAMP complex, a positive regulator of the lactose operon. (a) In the absence of large amounts of glucose, cAMP levels are high and the CRP–cAMP complex binds to a site near the promoter, where it activates transcription. (b) In the presence of large amounts of glucose, cAMP levels are low and the CRP–cAMP complex does not bind, so transcription is not induced to high levels, even in the presence of lactose.
The concentration of the small molecule cAMP in the cell is a signal about the nutritional state of the cell. In the absence of glucose, cAMP levels are high, and cAMP binds to CRP, changing the shape of CRP so that it can bind at a site near the operator and stimulate binding of RNA polymerase to transcribe lacZ and lacY when lactose is present in the cell (Fig. 19.19a). In this way, cAMP is an allosteric activator of CRP binding. However, if lactose is not present in the cell, the lactose repressor binds to the lactose operator and prevents transcription even in the presence of the cAMP–CRP complex (see Fig. 19.16).
In the presence of glucose, cAMP levels are low, and the cAMP–CRP complex does not bind the lactose operon. As a result, even in the presence of lactose, the lactose operon is not transcribed to high levels (Fig. 19.19b). In this way, E. coli preferentially utilizes glucose when both glucose and lactose are present and utilizes lactose only when glucose is depleted.