Cellular differentiation increasingly restricts alternative fates.
At each successive stage in development in which the cells differentiate, they lose the potential to develop into any kind of cell. The fertilized egg is totipotent because it can differentiate into both the inner cell mass and supporting membranes, and eventually into an entire organism. The cells of the inner cell mass, called embryonic stem cells, are pluripotent because they can give rise to any of the three germ layers, and therefore to any cell of the body. However, pluripotent cells cannot on their own give rise to an entire organism, as a totipotent cell can. Cells further along in differentiation are multipotent; these cells can form a limited number of types of specialized cell. Cells of the germ layers are multipotent because they can give rise only to the cell types specified for each germ layer in Figure 20.1. Totipotent, pluripotent, and multipotent cells are all stem cells, cells that are capable of differentiating into different cell types.
Quick Check 1 From what you know about embryonic development, do you think that a cell from the inner cell mass or one from the ectoderm has more developmental potential?
Quick Check 1 Answer
A cell from the inner cell mass of a blastocyst has more developmental potential than a cell from the ectoderm, which is one of the three germ layers, because the blastocyst occurs earlier in development than the formation of the three germ layers.
Why do differentiating cells increasingly lose their developmental potential? One hypothesis focuses on gene regulation. When cells become committed to a particular developmental pathway, genes no longer needed are turned off (that is, repressed) and are difficult to turn on again. Another hypothesis is genome reduction: As cells become differentiated, they delete the DNA for genes they no longer need.
These hypotheses can be distinguished by an experiment in which differentiated cells are reprogrammed to mimic earlier states. If loss of developmental potential is due to gene regulation, then differentiated cells could be reprogrammed to become pluripotent or multipotent. If loss of developmental potential is due to genome reduction, then differentiated cells could not be reprogrammed to become pluripotent or multipotent.
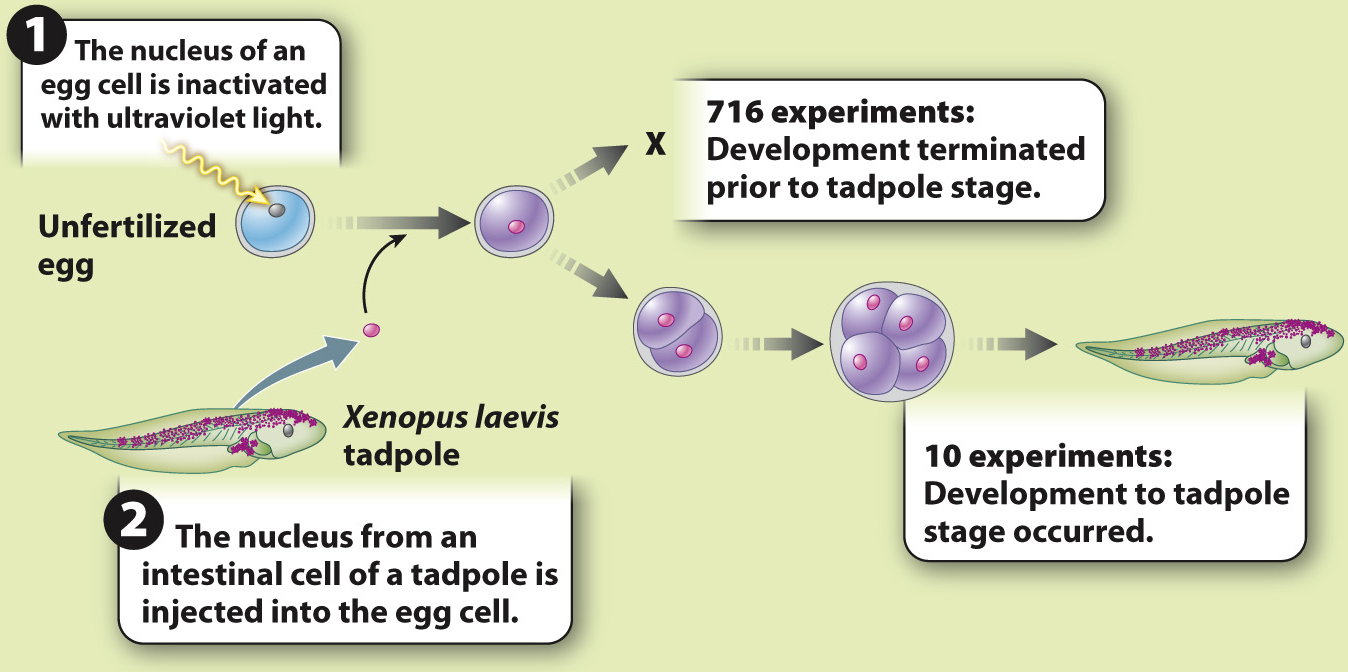
British developmental biologist John Gurdon carried out such experiments in the early 1960s (Fig. 20.2). Gurdon used a procedure called nuclear transfer, in which a hollow glass needle is used to insert the nucleus of a cell into the cytoplasm of an egg whose own nucleus has been destroyed or removed. Previous nuclear transfer experiments had been carried out in the leopard frog, Rana pipiens. Whereas nuclei from pluripotent or multipotent cells could often be reprogrammed to develop into normal tadpoles, attempts with nuclei from fully differentiated cells failed.
HOW DO WE KNOW?
FIG. 20.2
How do stem cells lose their ability to differentiate into any cell type?
BACKGROUND During differentiation, cells become progressively more specialized and restricted in their fates. Early studies left the mechanisms of differentiation unclear.
HYPOTHESIS One hypothesis is that differentiation occurs as a result of changes in gene expression. A second hypothesis is that differentiation occurs as a result of genome reduction, in which genes that are not needed are deleted.
EXPERIMENT John Gurdon carried out experiments in the amphibian Xenopus laevis to test these hypotheses. He transferred nuclei from differentiated cells into unfertilized eggs whose nuclei had been inactivated with ultraviolet light. If differentiation is due to changes in gene expression, then the differentiated nucleus should be able to reprogram itself in the egg cytoplasm and differentiate again into all the cells of a tadpole. If differentiation is accompanied by loss of genes, then differentiation is irreversible and development will not proceed.
RESULTS The experiment was carried out 726 times. In 716 cases, development did not occur; in 10, development proceeded normally.
CONCLUSION Although the experiment succeeded in only 10 of 726 attempts, it showed that the nucleus of an intestinal cell and the cytoplasm of the unfertilized egg are able to support complete development of a normal animal. This result allows us to reject the hypothesis that differentiation occurs by the loss of genes. The first hypothesis—
FOLLOW-
SOURCE Gurdon, J. B. 1962. “The Developmental Capacity of Nuclei Taken from Intestinal Epithelium Cells of Feeding Tadpoles.” Journal of Embryology & Experimental Morphology 10:622–
Gurdon tried the experiments in a different organism, the clawed toad Xenopus laevis, and demonstrated that nuclei from fully differentiated intestinal cells could be reprogrammed to support normal development of the tadpole (Fig. 20.2). Only 10 of 726 experiments succeeded, but this was sufficient to show that intestinal cell nuclei still contained a complete Xenopus genome. In other words, his findings supported the first hypothesis—
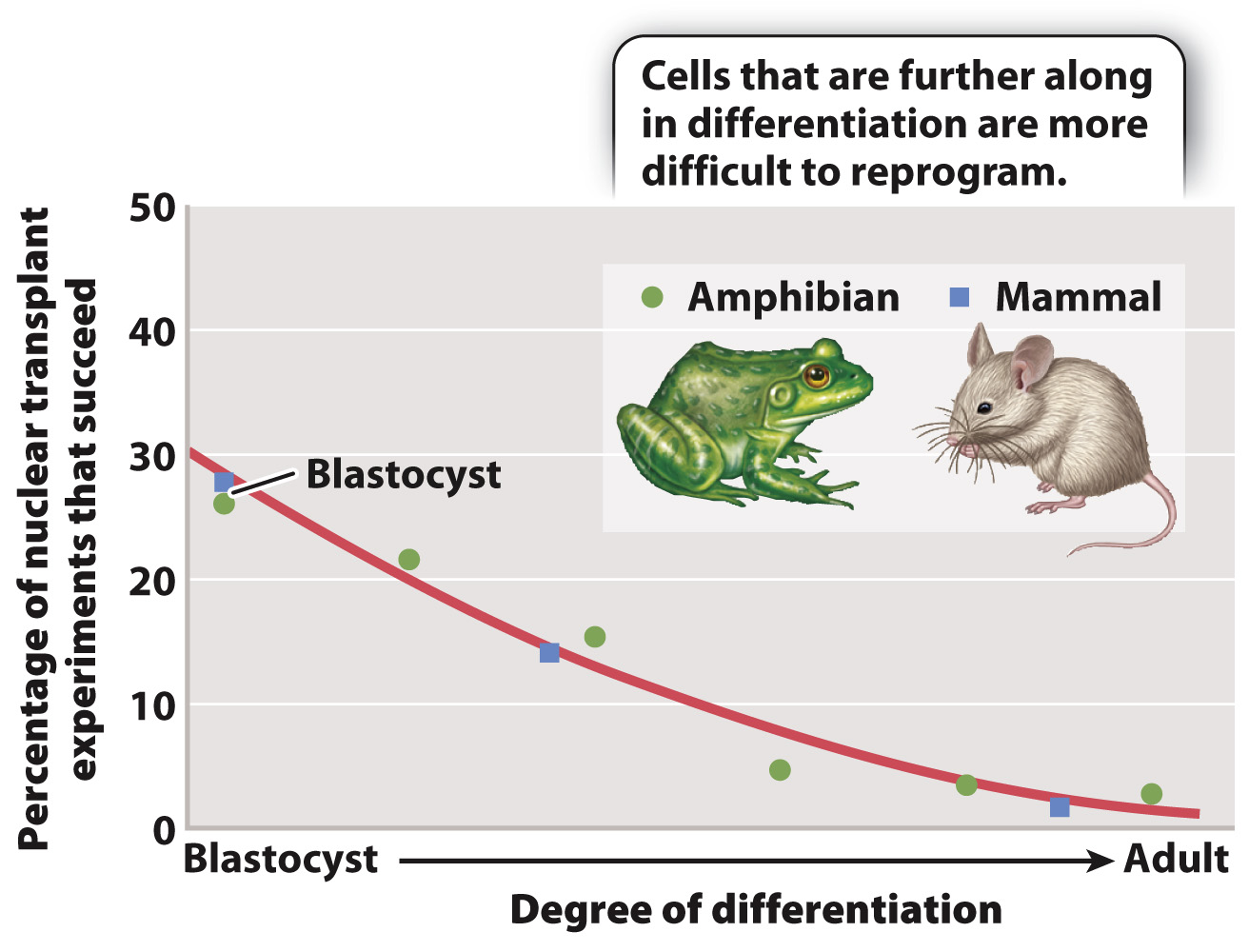
Fig. 20.3 summarizes the results of many nuclear transfer experiments carried out in mammals and amphibians. The percentage of reprogramming experiments that fail increases as cells differentiate. The best chance of success is to use pluripotent nuclei from cells in the blastocyst (or its amphibian equivalent, the blastula). However, even some experiments using nuclei from fully differentiated cells have been successful.
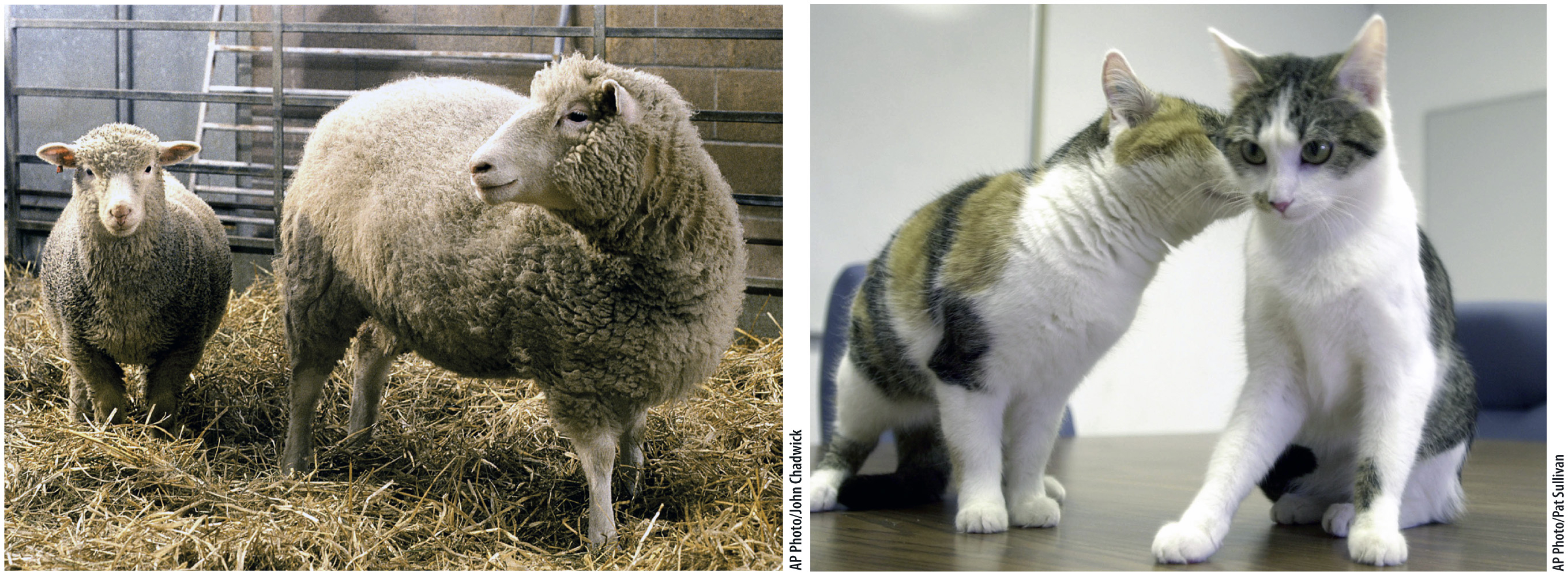
When nuclear transfer succeeds, the result is a clone—an individual that carries an exact copy of the nuclear genome of another individual. In this case, the new individual shares the same genome as that of the individual from which the donor nucleus was obtained. (The mitochondrial DNA is not from the nuclear donor, but from the donor of the egg cytoplasm.) The first mammalian clone was a lamb called Dolly (Fig. 20.4a), born in 1996. She was produced from the transfer of the nucleus of a cell in the mammary gland of a sheep to an egg cell with no nucleus, and was the only successful birth among 277 nuclear transfers. Successful cloning in sheep soon led to cloning in cattle, pigs, and goats.
402
The first household pet to be cloned was a kitten named CopyCat (Fig. 20.4b), born in 2001 and derived from the nucleus of a differentiated ovarian cell. CopyCat was the only success among 87 attempts. As shown in Fig. 20.4b, the cat from which the donor nucleus was obtained was a calico, but CopyCat herself was not, even though the two cats are clones of each other. The reason for their different appearance has to do with X-
403
Quick Check 2 X-
Quick Check 2 Answer
In Chapter 19, we saw that many common traits result from an interaction of genes and the environment, and so even clones can look different from each other because of environmental influences. Furthermore, if the clone is produced by nuclear transplantation, as in the cases of Dolly and CopyCat, the nuclear genomes of the clone and parent are identical, but their mitochondrial genomes are different.