Speciation can occur instantaneously.
Although speciation is typically a lengthy process, it can occasionally occur in a single generation, making it sympatric by definition. Typically, cases of such instantaneous speciation are caused by hybridization between two species in which the offspring are reproductively isolated from both parents.
For example, hybridization in the past between two sunflower species, Helianthus annuus and H. petiolaris (the ancestor of the cultivated sunflower) has apparently given rise to three new sunflower species, H. anomalus, H. paradoxus, and H. deserticola (Fig. 22.13). H. petiolaris and H. annuus have probably formed innumerable hybrids in nature, virtually all of them inviable. However, a few—

In many cases of hybridization, chromosome numbers may change. Two diploid parent species with 5 pairs of chromosomes, for a total of 10 chromosomes each, may produce a hybrid with double the number of chromosomes (that is, the hybrid inherits a full paired set of chromosomes from each parental species)—a total of 20. In this case, the hybrid has four genomes rather than the diploid number of two. We call such a double diploid a tetraploid. In general, animals cannot sustain this kind of expansion in chromosome complement, but plants are more likely to do so. As a result, the formation of new species through polyploidy—multiple chromosome sets (Chapter 13)—has been relatively common in plants.
Polyploids may be allopolyploids, meaning that they are produced from hybridization of two different species. For example, related species of Chrysanthemum appear, on the basis of their chromosome numbers, to be allopolyploids. Alternatively, polyploids may be autopolyploids, meaning that they are derived from an unusual reproductive event between members of a single species. In this case, through an error of meiosis in one or both parents in which homologous chromosomes fail to separate, a gamete may be produced that is not haploid. For example, Anemone rivularis, a plant in the buttercup family, has 16 chromosomes, and its close relative A. quinquefolia has 32. Anemone quinquefolia appears to be an autopolyploid derived from the joining of A. rivularis gametes with two full sets of chromosomes each.
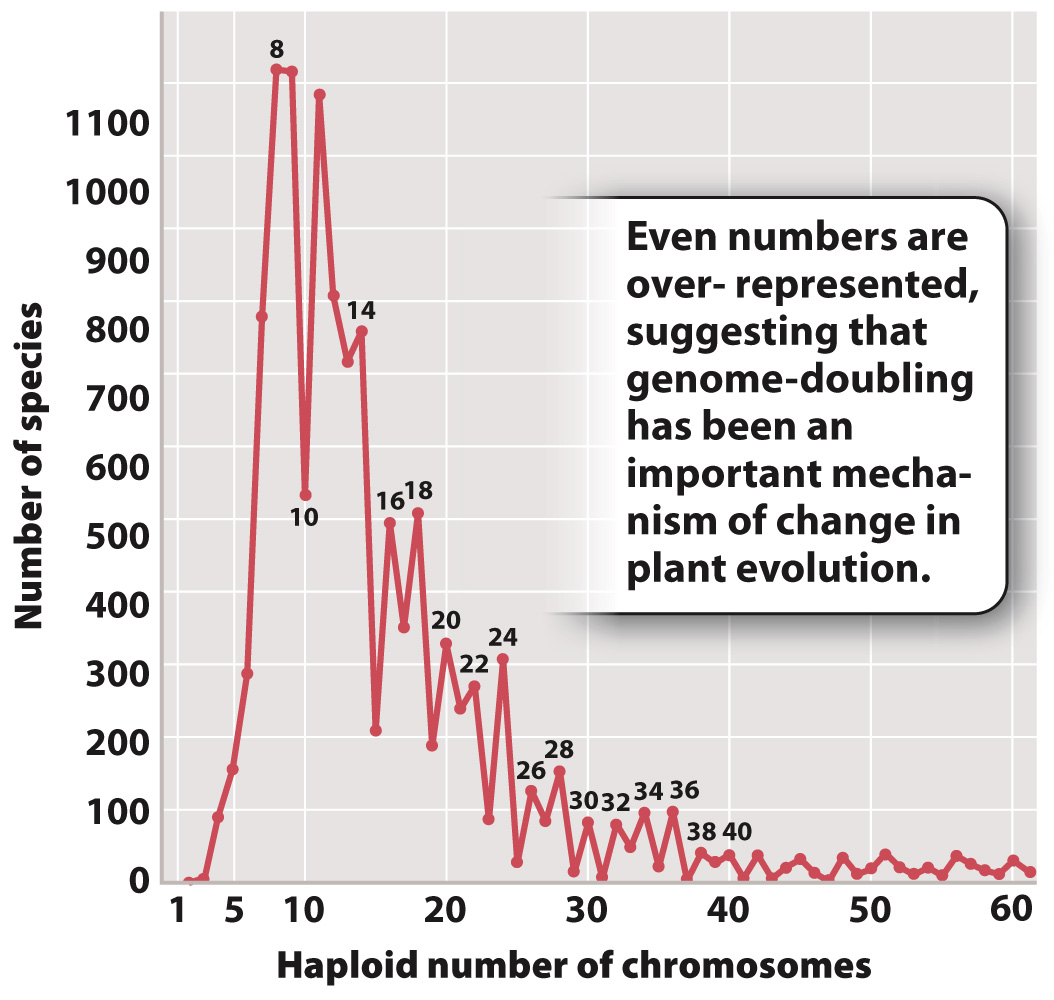
So rampant is speciation by polyploidy in plants that it affects the pattern of chromosome numbers across all plants. In Fig. 22.14, we see the haploid chromosome numbers of thousands of plant species plotted on a graph. Note that as the numbers get higher, even numbers tend to predominate, suggesting that the doubling of the total number of chromosomes (a form of polyploidy that will always result in an even number of chromosomes) is an important factor in plant evolution.
459