Archaeplastids, which include land plants, are photosynthetic organisms.
After the opisthokonts, the most conspicuous and diverse eukaryotes fall within the superkingdom Archaeplastida, another major branch of the eukaryotic tree (Fig. 27.14a). This is where we find the land plants, whose 400,000 described species dominate eukaryotic biomass on this planet (Chapter 33). The archaeplastids contain three major phylogenetic groups, and with the exception of a few parasitic and saprophytic (living on dead matter) species, all are photosynthetic. This indicates that the last common ancestor of living archaeplastids was photosynthetic. In fact, archaeplastids are the direct descendants of the protist that first evolved chloroplasts from endosymbiotic cyanobacteria. Some molecular analyses also place cryptophyte and haptophyte algae within this branch, but this remains a topic of continuing research.
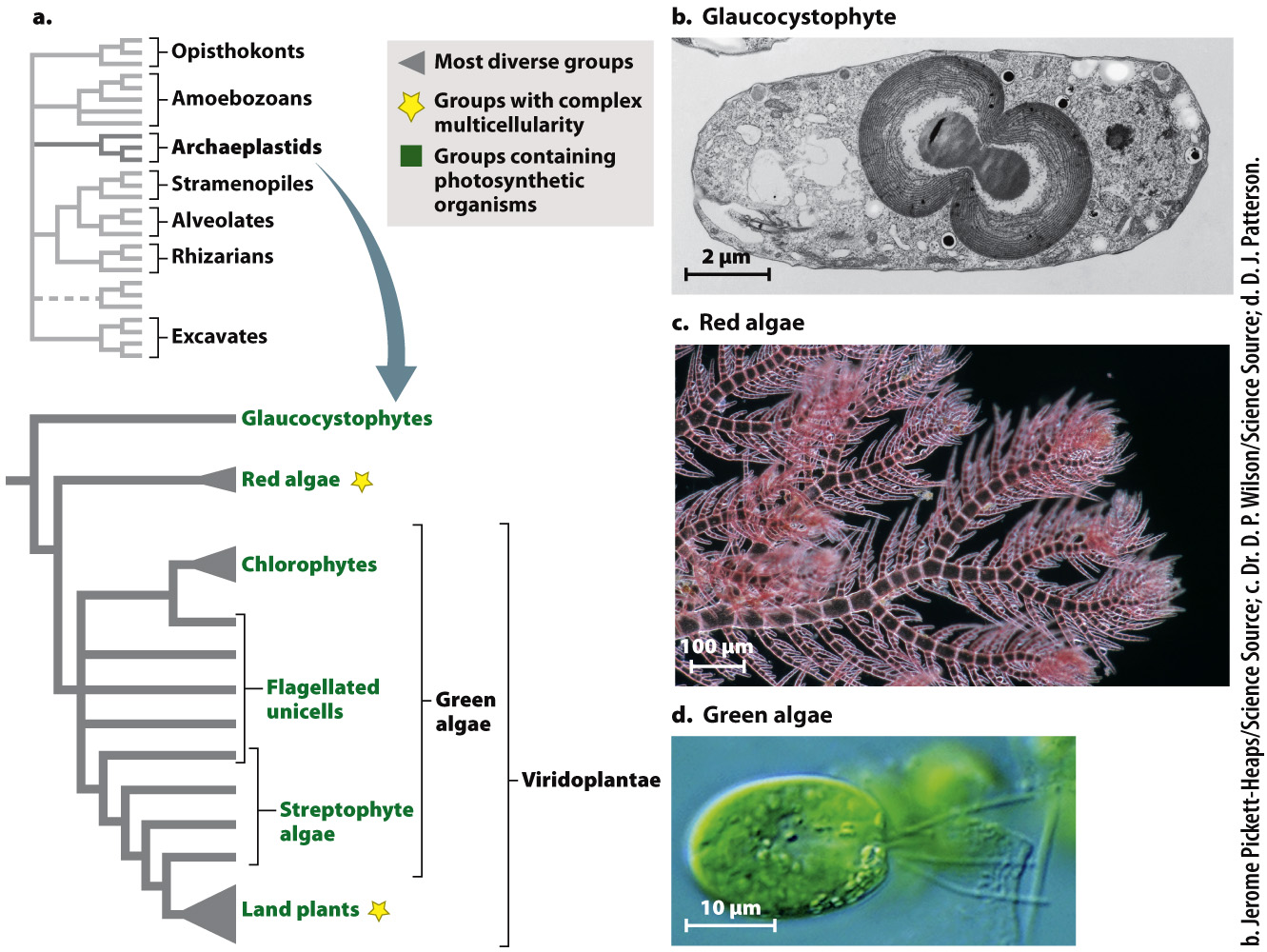
The least conspicuous archaeplastids are the glaucocystophytes (Fig. 27.14b), a small group of single-
More abundant and diverse is the second major archaeplastid group, the red algae (Fig. 27.14c). About 5000 species are known, mostly from marine environments. Most are multicellular and some have differentiated tissues and complex morphology. Like other archaeplastids, most have walls made of cellulose. The principal photosynthetic pigments in red algae are a form of chlorophyll called chlorophyll a and the biliproteins also found in glaucocystophytes and most cyanobacteria.
Red algae can be conspicuous as seaweeds on the shallow seafloor. One subgroup of red algae, known as the coralline algae, secretes skeletons of calcium carbonate (CaCO3) that deter predators and help the algae withstand the energy of waves in shallow marine environments. If you examine the margin of coral reefs, where waves crash into the reef, you will often see that it is coralline algae and not the corals themselves that resist breaking waves and so enable reefs to expand upward into wave-
Red algae are important as food in certain cultures, for example, as laver in the British Isles (particularly in Wales, in cakes similar to oatcakes) and as nori in Japan. Without knowing it, you often ingest carageenan, a polysaccharide synthesized by red algae. Carageenan molecules stabilize mixtures of biomolecules and promote gel formation, and so they are used in products ranging from ice cream to toothpaste. Agar, another red algal polysaccharide, is found universally in microbiology laboratories because the gel it forms at room temperature provides a nearly ideal substrate for bacterial growth.
The land plants and their algal relatives form Viridoplantae, the third branch of Archaeplastida. In this chapter, we are concerned with the protistan members of this group, the green algae (Fig. 27.14d). Green algae include a diverse assortment of species (about 10,000 have been described) that live in marine and, especially, freshwater environments. Green algae range from tiny single-

567
It is thought that green algae originated as small flagellated cells, and today most species that branch early on the green algal tree still live as photosynthetic cells in marine or fresh waters (Fig. 27.15a). The larger diversity of green algae, however, is found on two branches that diverged from these flagellated ancestors. One branch, the chlorophytes, radiated mostly in the sea and includes common seaweeds seen globally along seashores (Fig. 27.15e). Chlorophyte green algae have also played a major role in laboratory studies of photosynthesis (Chlamydomonas, Fig. 27.15a) and multicellularity (Volvox, Fig. 27.15d).
For humans, however, the more important branch of the green algal tree is the one that diversified in freshwater and, eventually, on land. Called streptophytes, the species on this branch show a progression of form from unicells at the base through simple cell clusters and filaments on intermediate branches (Figs. 27.15b and 27.15c), to complex multicellular algae that form the closest relatives of land plants (Chapter 33). As was true of choanoflagellates and animals, green algae display features of cell biology and genomics that tie them unequivocally to land plants.
568
Quick Check 2 Do plants and animals have a common multicellular ancestor?
Quick Check 2 Answer
No. Plants form one branch of the green algal tree, whose early branches are all occupied by unicellular forms. Similarly, the branch containing animals has choanoflagellates and other unicellular forms in its lower branches. Thus, animals and plants evolved multicellularity independently of each other. (Note that genetic evidence further supports this phylogenetic hypothesis: Plants and animals have different sets of genes that regulate multicellular development; see Chapter 28.)