Oxygen is necessary for complex multicellular life.
Earlier in the chapter, we discussed the key biological requirements for complex multicellularity. There is a critical environmental requirement as well: the presence of oxygen. There are other potential electron acceptors for respiration, like sulfate and ferric iron. However, they not only occur in much lower abundances than oxygen, but they are not gases and so do not accumulate in air. Only the oxidation of organic molecules by O2 provides sufficient energy to support biological communities that include large and active predators like wolves and lions. And only oxygen in concentrations approaching those of the present day can diffuse into the interior cells of large, active organisms. On our planet—
591
On the present-
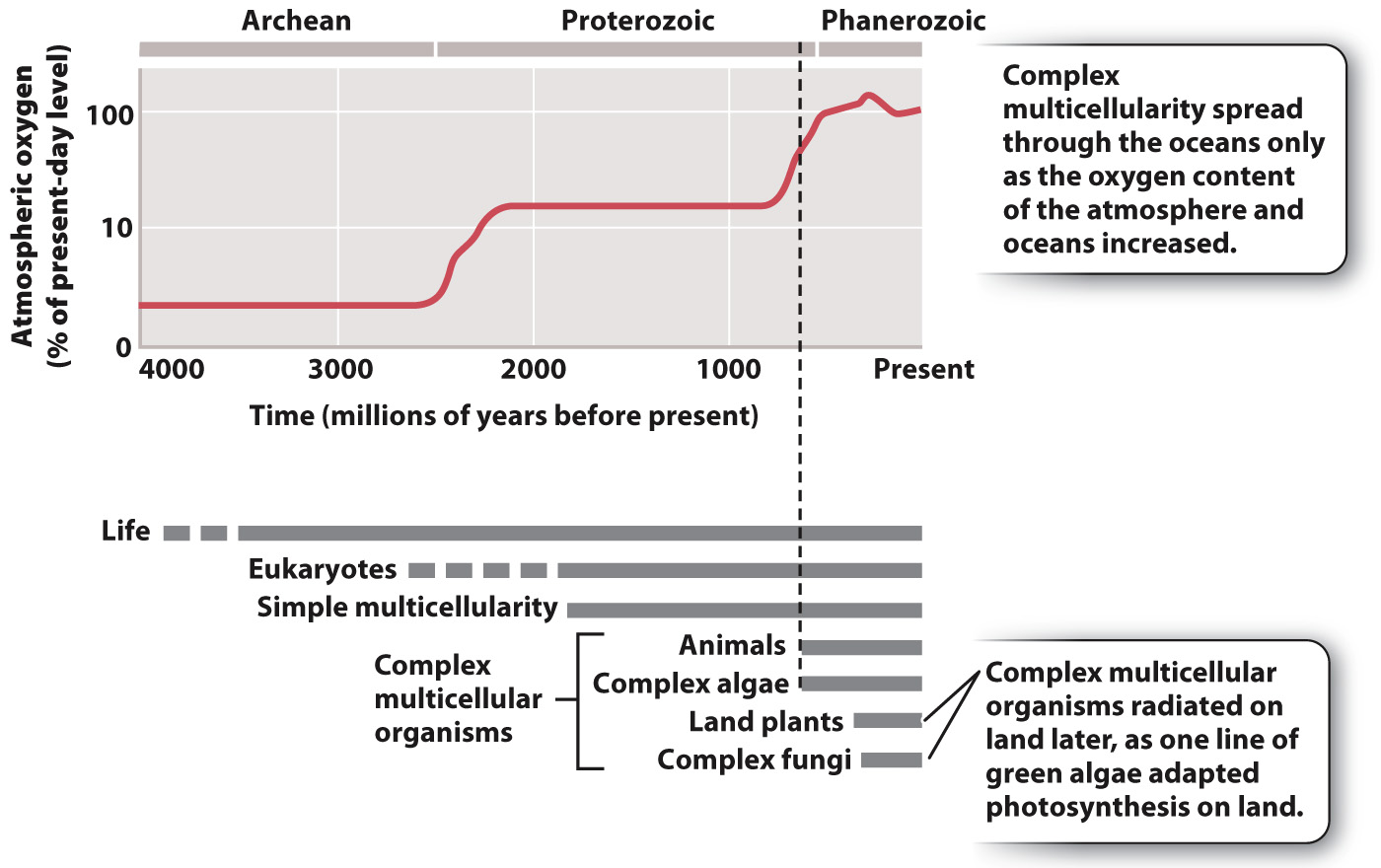
Scientists continue to debate the causes of this environmental transformation, but the correspondence in time between oxygen enrichment and the first appearance of large, complex animals (and algae) suggests that more oxygen permitted greater size and more energetic modes of life. Greater size in turn created opportunities for tissue differentiation, leading to transport of nutrients and signaling molecules by bulk flow. The evolution of bulk flow in turn permitted still larger size, setting up a positive feedback that eventually resulted in the complex marine organisms we see today.
With morphological complexity came new functions, including predation on other animals. Protozoan predators capture other microorganisms, but do so one cell at a time. In contrast, animals that obtain food by filtering seawater gather cells by the thousands, and larger animals can eat smaller ones, opening up endless possibilities for evolutionary specialization and, therefore, diversification. Among photosynthetic organisms, multicellular red and green algae were able to establish populations in wave-
The key point is that complex multicellular organisms didn’t succeed by doing the same things as simpler eukaryotes. Complex multicellularity spread through the oceans because it opened up new and unprecedented evolutionary possibilities.