The diversity of floral morphology is related to modes of pollination.
The evolutionary histories of the angiosperms and their animal pollinators are closely intertwined. A rapid increase in angiosperm diversity occurred between 100 and 65 million years ago, about the same time as the diversification of bees (Hymenoptera) and butterflies (Lepidoptera). The evolution of flowers allowed animals to specialize on new food resources. At the same time, animal pollinators greatly facilitated the movement of pollen between plants within the same population.
Flowers communicate their presence through both scent and color. For pollination to be reliable, however, it must be in the interest of the animal pollinator to move repeatedly between flowers of the same species. Flowers earn fidelity from their pollinators by providing rewards, frequently in the form of food (Fig. 30.14). Many insects consume some of the pollen itself, while many flowers also produce nectar, a sugar-


Some species provide rewards other than food. For example, many orchids secrete chemicals that male bees need to make their own sexual pheromones. Other flowers provide an enclosed and sometimes heated chamber in which insects can aggregate. But some flowers do not provide rewards at all: Instead, they “trick” their pollinators into visiting. For example, flowers that look and smell like rotting flesh attract flies, and orchids that look like a female bee even emit similar pheromones to attract male bees (Fig. 30.15). Male bees find these orchids irresistible and, as they attempt to copulate with the flower, deliver and receive pollen.
632
Producing rewards and attractants such as nectar and petals requires resources. Thus, while animals can provide more efficient pollination than can the wind, the resources spent may be wasted if nonpollinating visitors “steal” pollen or nectar. For this reason, many flowers have mechanisms to protect their investments. For example, tubular cardinal flowers provide rewards to hummingbirds able to probe their depths, and snapdragons provide rewards only to insects that are strong enough to push apart their petals.
Many plants have evolved flowers that match rewards and attractants to the metabolic needs and sensory capabilities of their pollinators. For example, flowers pollinated by larger pollinators such as bats and birds produce copious amounts of nectar. Bat-
HOW DO WE KNOW?
FIG. 30.16
Are pollinator shifts associated with the formation of new species?
BACKGROUND Columbines (Aquilegia species) are diverse and have visually striking flowers. Columbine flowers have nectar spurs, which are modified petals that form tubular outgrowths. Blue flowers with short nectar spurs are pollinated by bumblebees, red flowers with medium nectar spurs are pollinated by hummingbirds, and white or yellow flowers with long nectar spurs are pollinated by hawkmoths. The size of the nectar spur corresponds to the length of the tongue of the pollinator.
HYPOTHESIS The recent radiation of columbines corresponds with shifts from short-
METHOD Researchers mapped changes in flower color and nectar spur length onto a phylogenetic tree of columbines. To do this, they assigned each of 25 species examined to one of three groups based on length of nectar spur, flower color, and most common pollinator (bumblebees, hummingbirds, or hawkmoths). The group is indicated by the color circle at the tip of each branch in the phylogenetic tree in the figure. The researchers then did a statistical analysis that suggested the most likely pollinator type at each node representing an ancestor, also indicated by color circles in the figure. They used this information to infer the history of pollinator shifts during the radiation of this group.
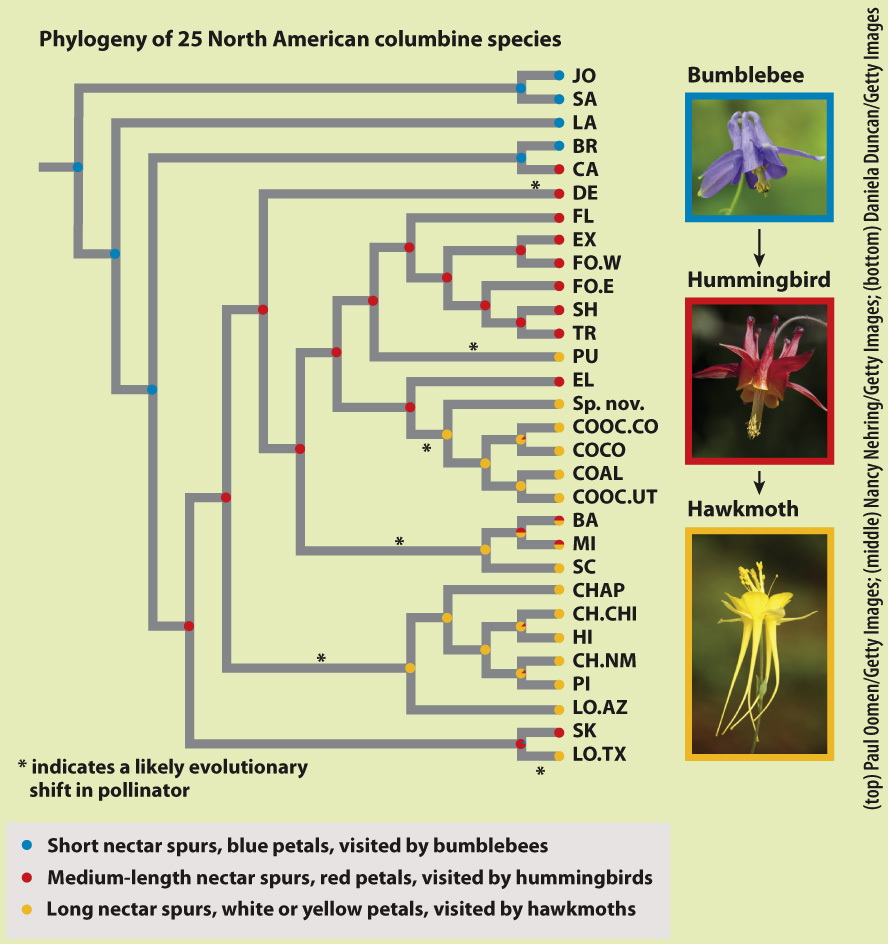
RESULTS Within the recent radiation of columbines, there are multiple instances of shifts from a shorter-
CONCLUSION The evolution of floral diversity and increasing nectar-
FOLLOW-
SOURCES Whittall, J. B., and S. A. Hodges. 2007. “Pollinator Shifts Drive Increasingly Long Nectar Spurs in Columbine Flowers.” Nature 447:706–
Angiosperms are thought to have evolved in tropical forests. Pollen cannot move freely through the air in the high density of foliage that is present year round in these forests, favoring the evolution of animal pollination. However, as angiosperms diversified and spread around the globe, some evolved wind pollination once again. The reversal to wind pollination is associated with habitats where pollinators are not reliably abundant, such as dry regions.
Wind pollination is most effective in species that are locally abundant. For example, many grasses rely on wind for pollination. Wind pollination is also common in temperate-