Bryophytes illustrate how the alternation of generations allows the dispersal of spores in the air.
In Chapter 29, we saw that bryophytes—
Like Chara and Coleochaete, Polytrichum has a photosynthetic body—
Their reliance on external water for fertilization helps explain why bryophytes produce their gametes near the ground, where they are most likely to encounter a continuous film of water. It also helps explain why bryophytes tend to be small, as sperm are able to swim only a couple of centimeters. Many bryophytes release their sperm only when agitated by raindrops. Raindrops signal the presence of surface water, and their impact can splash sperm farther than they could swim on their own.
622
It is after the fusion of egg and sperm gives rise to a diploid zygote that the life cycles of plants and their green algae relatives diverge. In plants, the zygote does not undergo meiosis, and it does not disperse. Instead, the zygote is retained within the female reproductive organ, where it divides repeatedly by mitosis to produce a new multicellular plant—
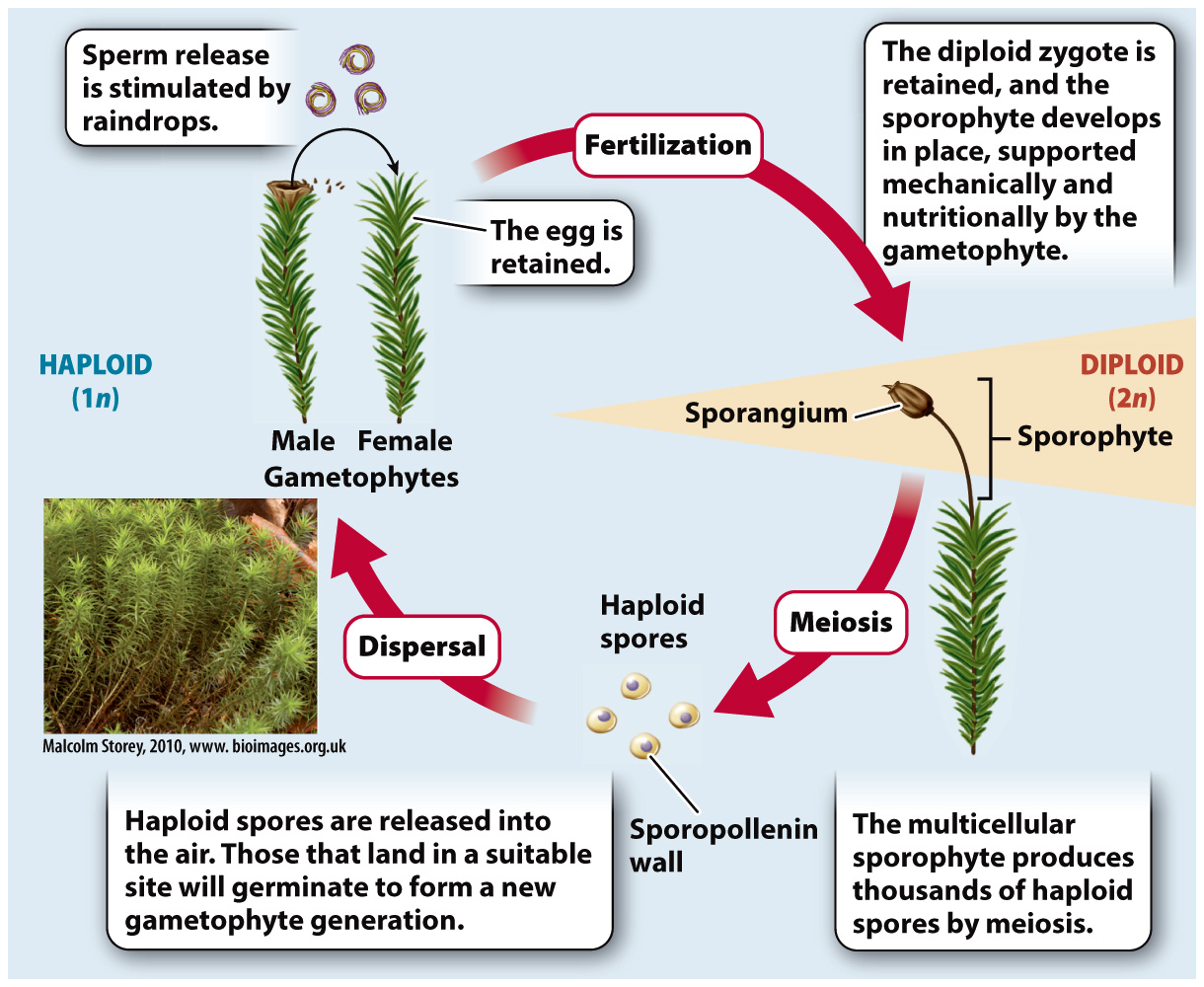
Because land plants are descended from green algae with only haploid gamete-
To answer this question, let’s look at the sporophyte of Polytrichum. Late in the growing season, the green tufts of this moss sprout an extension—

The significance of the multicellular sporophyte is that it enhances the ability of plants to disperse on land. The capsule at the top of the Polytrichum sporophyte is a sporangium, a structure in which many thousands of diploid cells undergo meiosis, producing huge numbers of haploid spores. In contrast, fertilization in Chara and Coleochaete is followed by meiosis in only a single cell (the zygote).
Spores are small and can be carried for thousands of kilometers by the wind. But their tiny size puts spores at risk of being washed from the air by raindrops. The sporangia of many bryophytes release their spores only when the air is dry. In Polytrichum, the sporangium has small openings from which it releases spores the way a salt shaker releases salt. When the air is wet, the spores form clumps that are too big to fit through these opening. More common is the presence of a ring of teethlike projections that curl over the top of the sporangia. These bend backward when air humidity is low, flinging the spores into the air (Fig. 30.5b).
What protects the spores from drying out or from exposure to damaging ultraviolet radiation as they travel through the air? Earlier, we noted that the zygotes of Chara and Coleochaete secrete a wall that protects the zygote within from desiccation. The wall contains sporopollenin, a complex mixture of polymers that is remarkably resistant to environmental stresses such as ultraviolet radiation and desiccation. In mosses and other land plants, it is the spore, not the zygote, that secretes sporopollenin. This suggests that early land plants retained a capacity present in their algal ancestors—
623
Following dispersal, most spores germinate within a few days, but some can survive within the spore wall for months or even years. When conditions for growth are favorable, the spore wall ruptures, and the haploid cell inside can then develop into a multicellular and free-